ピロリ菌による胃がん発症を抑制する酵素発見
─ EBウイルスの共感染では、酵素活性が弱められて胃がん形成が進む
畠山 昌則
2016年4月号掲載
ヒトの胃がんの80%以上は、ピロリ菌感染が原因となって発症する。ピロリ菌がどのように胃がんを引き起こすのか、そのメカニズム解明の研究で常に世界をリードしてきた畠山昌則教授(東京大学大学院医学研究科)。今回、ピロリ菌による胃がんの発症に抑制効果を持つ酵素SHP1が生体に備わっていることを発見。さらにその酵素活性は、EBウイルス(エプスタイン・バーウイルス)の共感染で低下することを明らかにし、Nature Microbiology に発表した。
―― ほとんどの胃がんの原因がピロリ菌感染によるとは、驚きです。
畠山氏: ピロリ菌(Helicobacter pylori)が胃がんと関係していることはよく知られています。多くの人は、ピロリ菌が引き起こすのは一部の胃がんのみと思っているのではないでしょうか。しかしながら、2014年のWHOの発表によると、全世界の胃がん患者の80%はピロリ菌感染が原因。とりわけ、日本をはじめ東アジアにおいては、98%以上、つまりほとんどの胃がんはピロリ菌感染が原因と考えられます。
―― 畠山先生は、ピロリ菌が胃がんを引き起こすメカニズム研究分野を切り開いてこられました。
畠山氏: ピロリ菌と胃がんの研究に私が着手したのは、17年ほど前のことです。当時一般に知られていたのは、ピロリ菌が産生するCagA(キャグエー)というタンパク質が何らかの役割を担っているかもしれないということだけ。それというのも、ピロリ菌にはCagAを産生する株と産生しない株の2種が存在するのですが(図1)、胃がん患者の胃から単離されるのはほぼすべてがCagA産生株だったからです。しかし、ピロリ菌が胃がん発症にかかわるメカニズムについては、当時は全くわかっていませんでした。
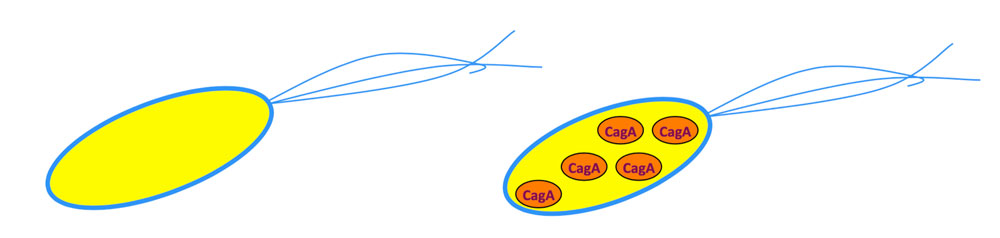
CagAの作用を解析
―― どのように研究を進められたのですか。
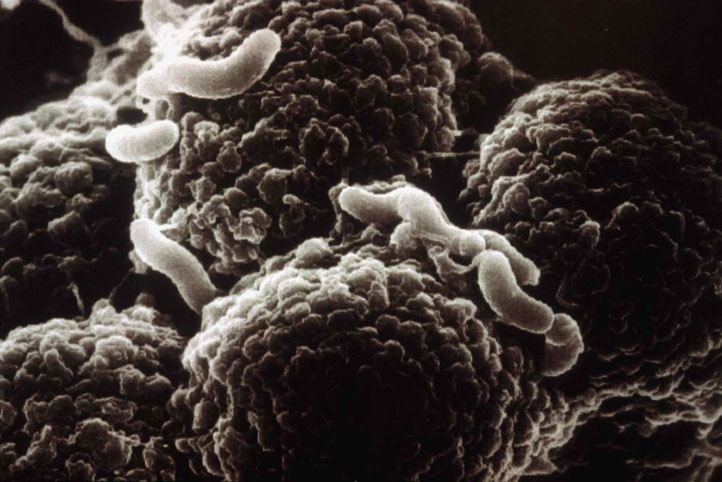
写真を拡大すると、胃壁の細胞に突起を差し込んでいるように見える。 | 拡大する
畠山氏: まず、ピロリ菌ゲノム配列の解析から、CagAタンパク質を作る遺伝子(cagA遺伝子)のそばに、おもしろい遺伝子群がクラスター(群れ)を作っていることがわかってきました。それらは、植物にがんを引き起こす細菌(アグロバクテリウム)が持つ遺伝子群の配列とよく似ていたのです。その細菌の遺伝子群は注射針のような働きを持つタンパク質構造体を作るのですが、胃がんを起こすピロリ菌CagA産生株も、体の表面からトゲ状の突起を出しています。そして、この突起を胃の壁(上皮細胞)に差し込んでいるように見えるのです。このトゲが注射針のように働き、CagAタンパク質を胃の細胞の中に注入しているのではないかと推察されました(図2)。
―― CagAは胃の細胞の中に打ち込まれる「発がんタンパク質」のようなものなのですね。
畠山氏: CagAタンパク質が胃の細胞の中で具体的にどのような効果を発揮するのか、それを突き止めるのにはずいぶん手こずりました。何の手がかりも見つからないまま1年間ほど苦戦していましたが、CagAタンパク質が胃の細胞の中でチロシンリン酸化という修飾を受けることが問題解決の突破口となりました。細菌のタンパク質であってもチロシンリン酸化を受けると、あたかもヒトタンパク質のように振る舞えるようになります。そこで、チロシンリン酸化を受けたCagAタンパク質が、ヒト細胞で重要な働き担う酵素などを誤作動させてしまうのではないかと考えました。
この仮説に沿って詳しく調べたところ、チロシンリン酸化されたCagAは、胃の細胞のSHP2という脱リン酸化酵素に特異的に結合することを突き止めました。その結果から、SHP2の酵素活性を異常に脱制御し暴走させることも明らかにしました。SHP2の暴走は最終的に、がん化につながる細胞の増殖異常や運動異常を引き起こすことがわかり、ピロリ菌と胃がんをつなぐ分子機構を解明することができたのです(図3)1。
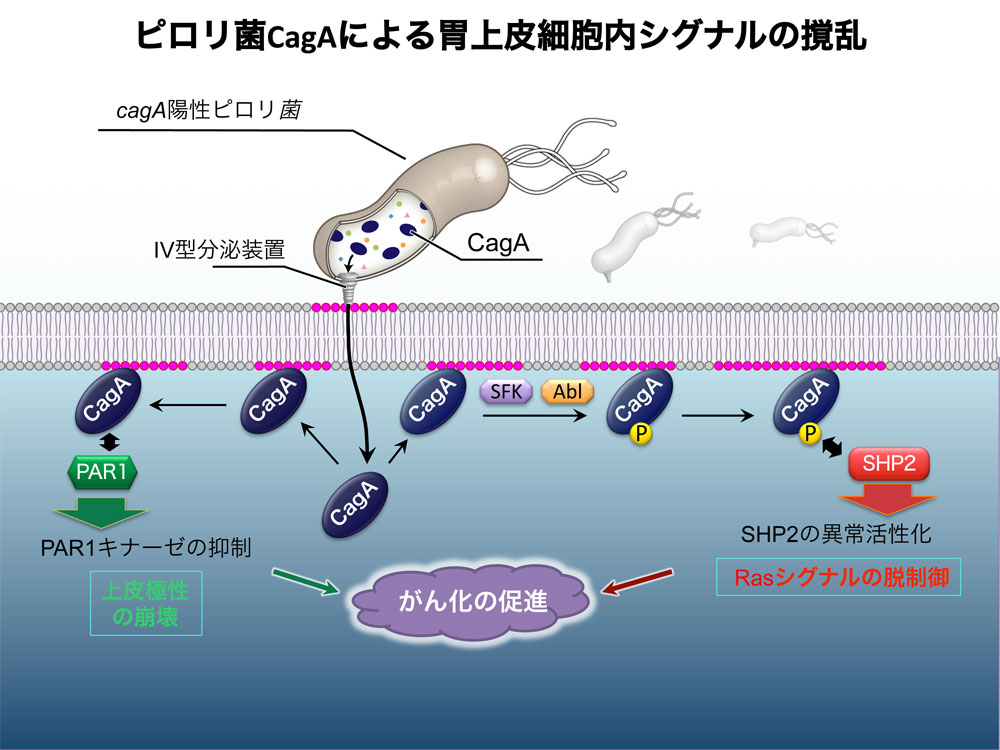
ピロリ菌のCagAタンパク質は、胃の上皮細胞内に注入され、2つの方法で発がんに関与する。
(a)まず、チロシンリン酸化されたCagAは、SHP2脱リン酸化酵素と結合してSHP2を異常に活性化し暴走させる。
(b)もう1つは、PAR1リン酸化酵素の働きを不活化して、細胞の極性を破壊する。 | 拡大する
―― CagAタンパク質の働きを次々と解明されたのですね。
畠山氏: さらにCagAタンパク質には、がんにつながる別の働きがあることも見つけました。胃の壁を形成する上皮細胞は、互いにコミュニケーションを取ることによって、一層の細胞層としてきちんと整列しています。つまり上皮細胞の極性(上下・前後・左右といった細胞内の位置取りにともなう機能の違い)が維持されています。ところがCagAタンパク質は、この細胞の極性を制御するPAR1というリン酸化酵素を不活化することがわかりました。その結果、上皮細胞の極性が破壊され、細胞にがん化しやすい状況が作られてしまうのです(図3)2。
マウスを用いた個体レベルでの実験でも「がんタンパク質」としてのCagAの働きを実証できました。遺伝子操作によりCagAタンパク質を全身で発現させたマウスを作ると、胃がんや小腸がん、血液がんの自然発症が促進することを確認できたのです。
また、CagAタンパク質の結晶化にも成功し、その構造解析からも、上記の発見を支持し、また発展させるようなデータが得られました3。
ピロリ菌CagAの働きを抑制する酵素を新たに発見
―― 今回の研究では、ピロリ菌のCagAの働きを抑制するヒトの酵素を見つけられたそうですね。
畠山氏: SHP2には、遺伝子配列がよく似た同じく脱リン酸化酵素の兄弟分子、SHP1が存在します。しかし、SHP1は主に血液系の細胞においてのみ発現すると報告されていたので、私たちはこれまで特に注目していませんでした。
あるとき、ピロリ菌と血液系のがんについて調べることになり、SHP1の発現を調べてみました。すると偶然にも、SHP1が胃の細胞でも発現していることを発見。驚きましたが、とりあえず胃でのSHP1の解析も進めてみることにしました。当初は、SHP1とSHP2は兄弟分子なので、同じような働きをすると考えられ、あまりおもしろい発見は期待できないだろうとも思っていました。ところが、予想は完全に裏切られました。意外なデータが出てきたのです。胃がんに対して、SHP1とSHP2は全く逆の働きをしていたのでした。
―― CagAに対してSHP1はどのような仕組みで働くのですか。
畠山氏: 一言でいうと、CagAタンパク質は胃細胞内でチロシンリン酸化を受けるのですが、SHP1は、そのCagAを脱リン酸化する(リン酸基を奪い取る)のです。すなわちSHP1は、CagAのチロシンリン酸化を抑制するように働き、その結果、CagAによるSHP2の異常な活性化が抑えられることになります(図4)。

SHP2とよく似た構造のSHP1は、CagAタンパク質からリン酸基(Pで表されている)を奪い取る働きをするので、CagAの結合によるSHP2の異常活性化・がん化が抑制される。 | 拡大する
―― 正反対の働きをするSHP2とSHP1ですが、そのバランスはどうなっているのでしょうか。
畠山氏: SHP1はピロリ菌の胃がん発がんに抑制的な効果を発揮しますが、胃の細胞の中では、SHP2の方が量的にSHP1を圧倒しているため、SHP1がピロリ菌CagAによる胃がんの発症にストップはかけられないのだろうと考えています。
SHP1の活性を人為的に上げることができれば、ピロリ菌による胃がんを抑えられる可能性もでてくるでしょう。胃がん治療の分子標的になることも期待されます。
EBウイルスの共感染がSHP1の活性を抑制
―― 逆に、SHP1の活性が弱まると、胃がんは発症しやすくなるのでしょうか。
畠山氏: そうだと考えられます。そこで、SHP1の活性を弱める実例がないかと探すことにしました。
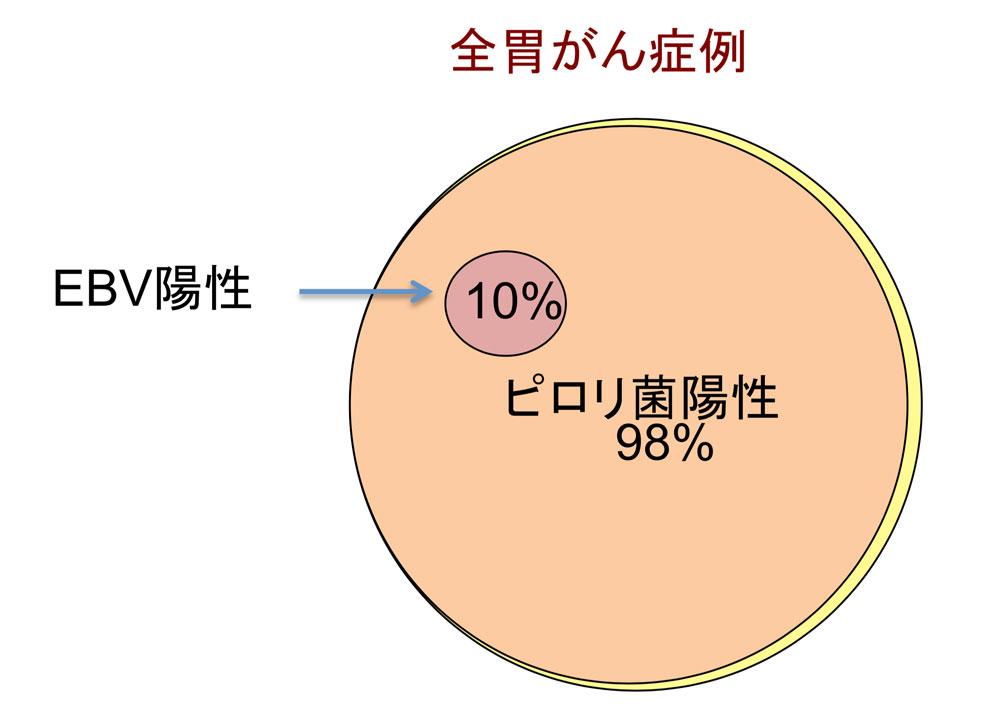
昔から胃がんに関係するといわれていたウイルスに、EBウイルス(エプスタイン・バーウイルス)があります。日本人の胃がんのほとんどがピロリ菌感染を原因としていると言いましたが、そのうちの約10 %の患者さんは、EBウイルスにも感染していることがわかっていました(図5)。さらに、EBウイルスに感染した胃がん細胞のゲノムを調べると、DNAの一部に高度なメチル化が起こっていることも知られていました。一般にDNAのメチル化は、そこに存在する遺伝子の発現が抑制されることを意味します。
そこで、もしEBウイルスがSHP1タンパク質を作る遺伝子の発現を抑制するとしたら、CagAの発がん活性が上がるだろうと考えたのです。実験の結果は予想通りでした。EBウイルスによるDNAメチル化が起こってSHP1遺伝子の発現が抑制され、CagAの発がん活性が上昇、その結果がん形成が進むという反応経路があることを実証できました。これが今回の研究の重要なポイントです。細菌とウイルスが発がんで連携し合うという、初めての報告になります。
―― Nature Microbiology に投稿することにしたきっかけは?
畠山氏: Nature Microbiology の創刊は、その1年ほど前にエディターの方からメールをいただいて知っていました。微生物の分野で第一級のジャーナルを作りたい、ついては、ぜひよい論文を投稿してくださいとのことでした。それで、今回、この論文を投稿することにしたのです。
今後に向けて
―― 日本には胃がんが多いので、なおさら研究に力が入りますね。
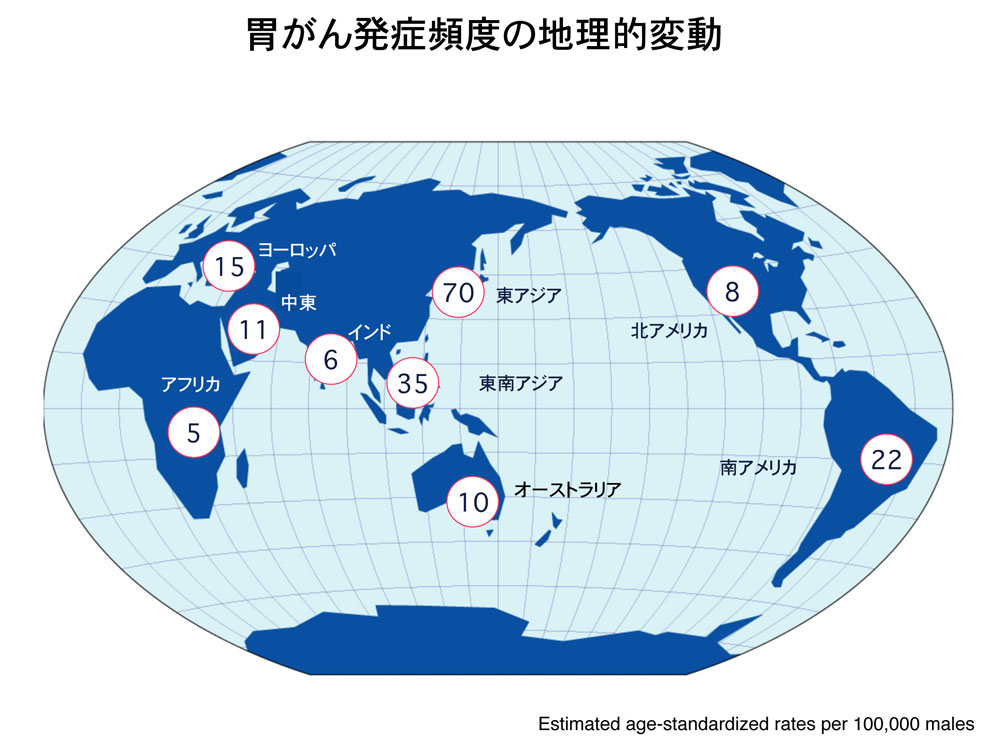
畠山氏: 胃がんの発症頻度は、東アジア(日本、中国、韓国)で圧倒的に多いのが特徴です。おそらく、世界のどの地域でも人口の半分以上がピロリ菌に感染しているにもかかわらず、世界の胃がんの60%以上が東アジアで起きているのです(図6)。東アジアと欧米でこんなにも胃がんの発症頻度が異なるのは、欧米ではCagAを産生しないピロリ菌株が多いからです。一方、日本では、ほとんどがCagA産生株です。
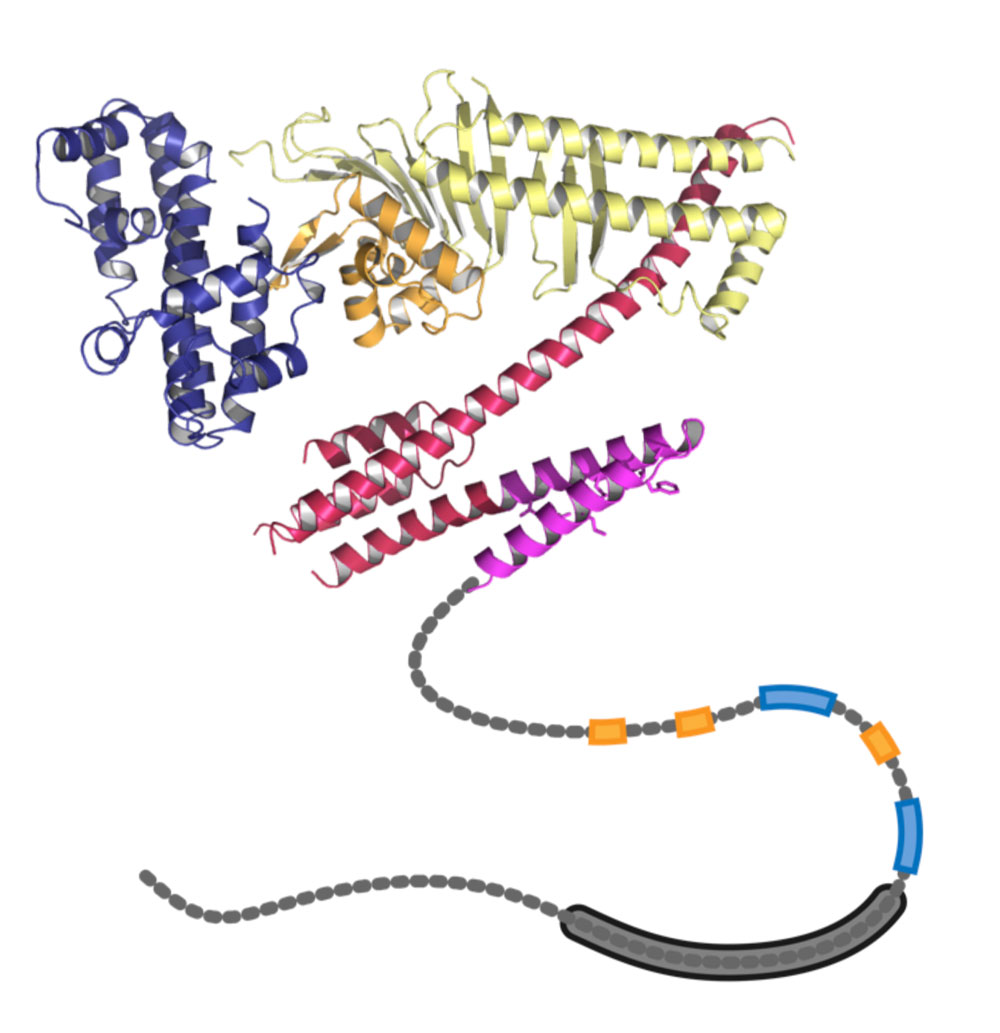
CagAの尾部は鞭のようにフレキシブルな構造をとっており、この構造を用いてSHP2と結合する。SHP2との結合にかかわる部位は東アジア型 CagAと欧米型 CagA間で形が異なり、前者は後者に比べてより強くSHP2と結合する。 | 拡大する
また、昨年私たちは、ピロリ菌から作られるCagAタンパク質の構造そのものが、欧米と東アジアで少し異なっていることを報告しています(図7)4。欧米型ではチロシンリン酸化CagAタンパク質とSHP2との結合力が弱く、SHP2を活性化しにくいのです。
―― 日本人にとって、ピロリ菌対処の重要性が明確ですね。
畠山氏: ピロリ菌は、人類誕生後間もなく人類に感染したと思われます。しかし、人類が現在のように長生きできなかった頃は、ピロリ菌は人類に「胃がん」のような悪さをしてこなかったでしょうし、狩猟中心であった欧米人に比較して菜食中心の生活をしてきた東アジア人には、胃酸を抑えるといったよい効果をもたらしてきた可能性もあるでしょう。しかしながら、長寿社会を迎え、ピロリ菌が胃がん発症の最大のリスク因子になってしまいました。
胃がんはまだまだ死亡率の高い、恐ろしいがんです。ピロリ菌は胃がんの原因として、十分条件ではありませんが、必要条件です。日本では人口の約半数である5,000〜6,000万人がピロリ菌に感染していると考えられており、この状況を放置しておけば、今後20〜30年間に約500万人が胃がんを発症すると推計されます。ピロリ菌の除菌はもちろん重要ですが、SHP1の活性を上げる方法などの研究にも力を注いでいく必要があると思います。
参考文献
- Higashi, H. et al., Science, 295: 683-6 (2002)
- Saadat, I. et al., Nature, 447: 330-333 (2007)
- Hayashi, T. et al., Cell Host Microbe, 12: 20-33 (2012)
- Nagase, L. et al., Sci. Rep., 5: 15749 (2015)
インタビューを終えて
ピロリ菌の作るCagAタンパク質の研究を15年以上も続け、先頭に立ってこの分野を開拓してきた畠山教授。「こんなに研究のしがいがある分子に出会うことは、なかなかないと思いますね。多機能で謎が尽きず、追いかければ追いかけるほど、いろいろな発見がある」。頑張れる動機は、「おもしろい」と思う気持ちとのこと。若手の指導にあたっても「本人が心からおもしろい」と思える研究テーマに出会う環境作りを心がけている。
藤川良子(サイエンスライター)
Nature Microbiology 掲載論文
Letter: 宿主SHP1ホスファターゼはピロリ菌(Helicobacter pylori)CagAの発がん活性に対抗するが、その発現はエプスタイン・バーウイルスにより抑制されうる
Nature Microbiology 1 : 16026 doi:10.1038/nmicrobiol.2016.26 | Published online 14 March 2016
Author Profile
畠山 昌則(はたけやま まさのり)
東京大学大学院医学系研究科医学部 病因・病理学専攻 微生物学講座 微生物学分野 教授
| 1981年 | 北海道大学医学部卒業 |
| 1986年 | 北海道大学大学院医学研究科博士課程修了、大阪大学細胞工学センター助手(谷口維紹教授) |
| 1991年 | 米国マサチューセッツ工科大学ホワイトヘッド研究所(Weinberg研究室)でポスドクとして研究に従事 |
| 1995年 | (財)癌研究会癌研究所ウイルス腫瘍部 部長 |
| 2000年 | 北海道大学遺伝子病制御研究所病態研究部門分子腫瘍分野 教授 |
| 2009年 | 現職 |
