植物は自分と他者を区別する
─ 他者の花粉を選んで受精するペチュニアの花
高山 誠司、久保 健一
2015年1月号掲載
Nature Plants が2015年1月に創刊された。その表紙を飾ったのは美しいペチュニアの花。ペチュニアは受精に際して、自分の花の花粉と、他の花の花粉を識別し、後者のみを受け入れる。近親交配を避けるこの巧みな仕組みを解明し、記念すべき創刊号に研究成果を発表した高山教授と久保研究員にお話しを伺った。
―― 植物は、「自分」と「他者」を区別している。驚きです。

雌しべの先端(乳頭細胞)に付いた自己の花粉と非自己の花粉を観察。非自己の花粉は約30分後から吸水を始めて、膨らんでくる。 | 拡大する
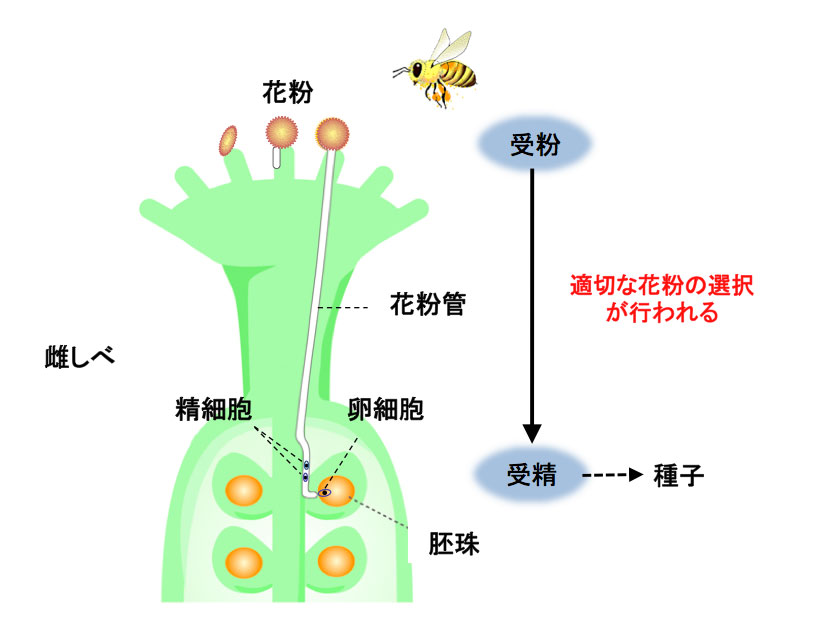
雌しべの先端に非自己の花粉が付くと、吸水・発芽して、花粉管が胚珠にまで伸びていく。花粉の精細胞が花粉管を通り、胚珠の卵細胞と受精し、やがて種子ができる。 | 拡大する
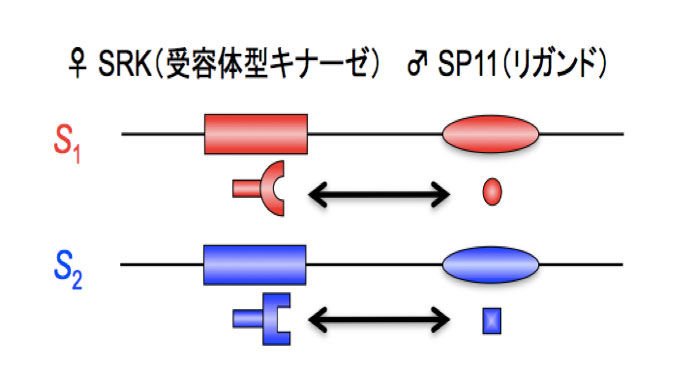
雌しべの酵素タンパク質(受容体型キナーゼ:SRKと呼ばれる)と、花粉のタンパク質(SP11と呼ばれる)が並んでコードされており、この並び方が保たれたまま遺伝していく。S1遺伝子のSRKは、同じS1遺伝子のSp11と結合することができる。 | 拡大する
高山氏: このアブラナの写真を見てください(図1)。同じアブラナでも、非自己(他の植物体)の花粉が雌しべに付いたときにだけ、変化が起こります。
非自己の花粉は水を吸って膨らみはじめ、花粉管が下方に伸びていきます。そして、花粉に含まれていた精細胞が、花粉管を通って雌しべの根元にまで運ばれ、卵細胞と受精して種子が作られます(図2)。
多くの植物は、自己の花粉を排除する性質(自家不和合性)を持っています。近親交配が起こると遺伝情報の多様性が失われていきますが、それを避けるような性質が進化してきたと考えられます。花粉を識別するのにどのような仕組みが働いているのかは謎で、人々は長い間不思議に思っていました。ダーウィンもその一人だったようですが、最近研究が進み、ようやく分子レベルの仕組みが明らかになってきました。
S遺伝子が自己と非自己の認識を決める
―― これまでの研究で、どのようなことが明らかになってきたのでしょうか。
高山氏: 自己と非自己の認識は、S遺伝子と呼ばれる遺伝子の働きによって生じることがわかっています。S遺伝子と名付けられたのはかなり昔のことで、実際には雌しべのタンパク質と、花粉のタンパク質が並んでコードされていています(専門用語ではSハプロタイプと呼ばれる)。このS遺伝子には、塩基配列の異なるバリエーション(S1、S2……Sn)が何種類も存在します。そして、花粉と雌しべが出会ったときに、花粉と雌しべが同じS遺伝子を持っていれば自己と認識することがわかりました。同じS遺伝子にコードされた花粉と雌しべのタンパク質はピタッと結合し、そこで花粉を排除するシグナルが発せられるため、花粉が受精できなくなるからです1(図3)。
―― アブラナ以外の植物でも、同じ仕組みが働くのですか。
高山氏: 植物の種類によっていろいろ異なることがわかってきています。私たちはナス科の園芸植物、ペチュニアについて調べました。そして、認識に関わる分子を見出し、アブラナが「自己を認識して、それを排除する」のに対して、ペチュニアは「非自己を認識して、それを受け入れる」という仮説を立てて、2003年に発表しました。ところが最初、このペチュニアの仮説の評判がとても悪かったのです。自己は1つだから見つけるのは簡単だが、非自己はたくさんの種類が存在するので、そんなことができる複雑な仕組みがあるわけないと。
久保氏: そこで、この仮説の証明が私の研究テーマになりました。
ペチュニアの仮説を2010年に証明
―― 高山先生の立てた仮説について、もう少し具体的に説明してください。
久保氏: はい。ペチュニアのS遺伝子からは、花粉を殺す「毒」と、非自己の「毒」を壊すことのできる「解毒剤」が作られるのではないかと考えられました。雌しべタンパク質が、毒(RNAを壊すRNA分解酵素)で、花粉タンパク質が、解毒剤(タンパク質を分解するF-box因子)をコードしていたからです。つまり、受精が起きるためには、雌しべがもつ毒を非自己の花粉がもつ解毒剤で解毒しなければならない。そのためには非自己を認識しているはずだと考えたのです。
―― 非自己を見つける仕組みをどのように証明したのですか。
久保氏: この仮説が成り立つためには、たくさんの毒に対処できるように、1個の花粉がいろいろな解毒剤を同時に持つことしか考えられません。実際にペチュニアのS遺伝子を詳しく調べると、1個の花粉が持つ花粉タンパク質は1つではなく、少しずつ異なるタンパク質が複数並んでS遺伝子にコードされていました。花粉タンパク質(F-box因子:SLFと表す)を、SLF1、SLF2、SLF3……と分類し、どの解毒剤がどの毒を解毒するか、1つずつ組み合わせてみて、検証することにしました。
毒と解毒剤をいろいろな組み合わせで持つペチュニアを遺伝子組み換え技術で作製し、解毒が起こるかどうかを実際に調べたのです。解毒は、花粉を雌しべに付けて、種子が出来れば解毒が起こっている、という具合に判断しました。地道な実験を繰り返し、1つの花粉が少なくとも6個の解毒剤を同時に持つこと、自分に対する解毒剤は持たないことを証明することができたのです。2010年に論文発表し、仮説が受け入れてもらえました2。
ペチュニアの仕組みの全貌を探る研究に着手
―― 今回の研究3は、このペチュニアの研究をさらに発展させたのですね。
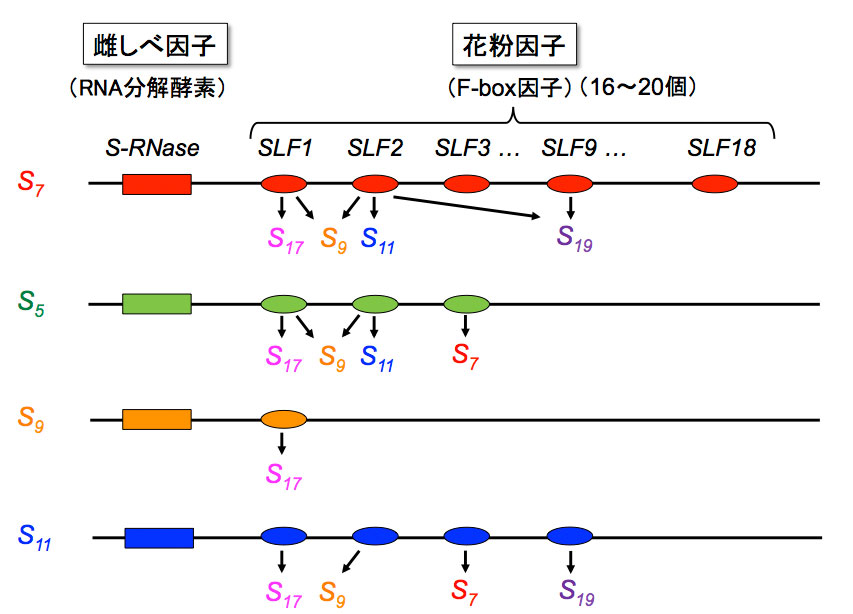
ペチュニア(ナス科)のS遺伝子には、RNA分解酵素(「毒」)1個と、F-box因子(「解毒剤」)16〜20個(18種類)が並んでコードされている。各解毒剤が数種類の毒を解毒し、このような解毒剤の協調的な働きにより、非自己を認識するとするモデル。 | 拡大する
高山氏: はい。自分以外の非自己をすべて認識して解毒するのに、6個の解毒剤ではいくらなんでも少なすぎると思ったのです。ですから、全部でいくつ持つのだろうか徹底的に調べようと思いました。結論からいうと、全部で16〜20個(種類としては18種類)の解毒剤を持つことがわかりました。各解毒剤は、それぞれ数種類ずつの毒を解毒することも明らかになりました。1つの花粉がこうした18種類の解毒剤を持つことによって、非自己を認識するということです(図4)。
―― 18種類で足りるのでしょうか。
久保氏: 先の研究と同じように、毒と解毒剤のいろいろな組み合わせの植物体を遺伝子組み換え技術で作成していくうちに、1つの解毒剤が何パーセント位の非自己の毒を解毒できるかがわかってきました。そして、これらの実験データをもとに統計的に計算してみると、40種類以上あることが知られる毒タンパク質を、18種類の解毒剤でほぼ識別可能という結果が得られました。
―― 系統樹も作成されたのですね。
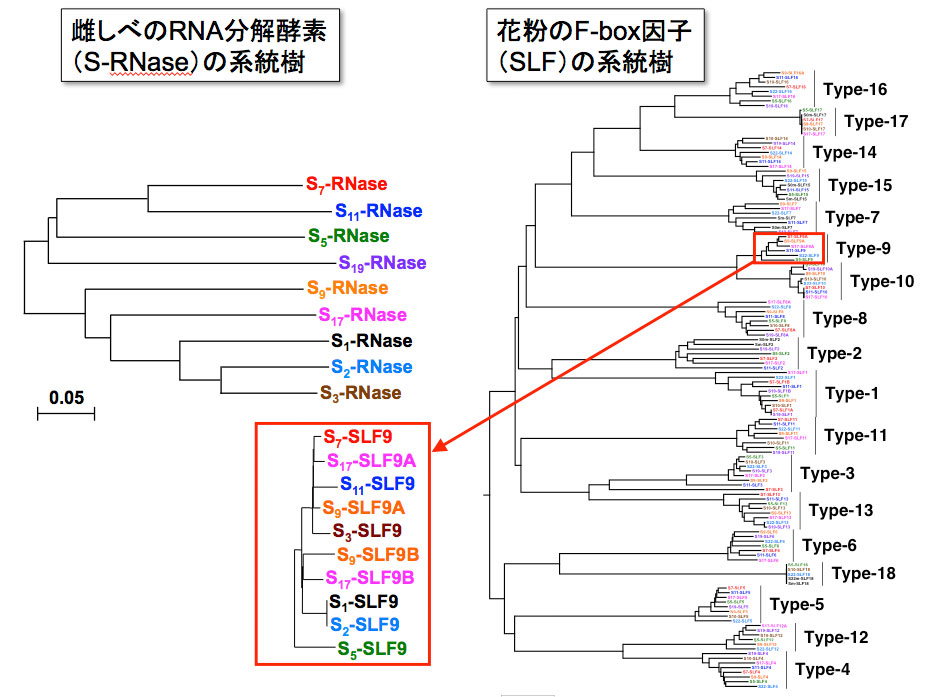
左側は毒(RNA分解酵素:S-RNase)、右側は解毒剤(F-box因子:SLF)の系統樹。横軸は時間。毒と解毒剤のタイプの組み合わせから、新たな毒が生じると、それに対応する解毒剤が生じることが推定された。 | 拡大する
高山氏: このような巧妙な仕組みをどのようにして持つに至ったか、進化の道筋についても調べたいと思いました。S遺伝子の配列を詳細に比較解析すると、それらの配列がどのように生成されてきたのかを示す系統樹を描くことができます。そこで、毒と解毒剤の遺伝子の塩基配列を詳細に比較して系統樹を作りました(図5)。
言葉では説明が難しいのですが、この系統樹から読み取れることは、ペチュニアの集団の中で、新たな毒を持つ個体が偶然生じると、それに対応する解毒剤が生じてくるということです。そして、効果的な解毒剤が生じると、その解毒剤を持つ個体は生存に有利なので子孫を残しやすく、その解毒剤が集団のなかで広まるということが推測できました。このような過程を引き起こす分子レベルの仕組みとしては、似た塩基配列がゲノム中に増える現象(遺伝子重複)や、他の個体に移動する現象(遺伝子交換)が考えられます。
―― 塩基配列の解析などの膨大な作業をご自身でやられたのですか。
久保氏: 次世代シーケンサーで行う作業は、スイス・チューリッヒ大学の清水健太郎教授にお願いしました。シーケンサーで得られたデータの解析や系統樹作成は、独学ですが、自分でやりました。大変な量のデータなので、地道にこつこつという言葉がまさしくぴったりの作業でした。遺伝子組み換え植物体の作成も数百植物体に及んだため、大変でしたけどね。私たちの研究を追ってくるライバルの存在も感じていたので、それも気にしつつ、がんばりました。
論文発表直前に、共同研究者がライバルに
―― ライバルとの激しい競争が?
久保氏: 実は、共同研究者に驚かされました。以前から共同研究者だった米国のグループが、「今回は自分たちも、独自に論文発表する」と連絡してきたのです。私たちは、論文を投稿する直前の段階。解析していたS遺伝子の種類は彼らより私たちのほうがかなり多く、議論できる視点も多いことはわかっていたのですが、それでもとても大きなプレッシャーとなりました。
そして、彼らは私たちより一歩先にPlant Cell に投稿。私たちが論文をリバイスしている段階で、彼らの論文が掲載されました。彼らは2種類のS遺伝子について解毒剤を解析し、私たちより1つ少ない17種類の解毒剤を同定していました。彼らは紳士的に事前に連絡をくれ、毒と解毒剤の命名法(番号付け)を私たちに合わせてくれていましたが、そのとき初めて論文の内容を知りました。
―― まあ、それは、さぞかし驚かれたことでしょう。高山先生たちは、最初からNature Plants への投稿を決めていたのですか。
久保氏: それが、違うのです。最初は、Nature Genetics に投稿しました。今回の研究は、遺伝学的な解析や分子系統学的な解析が主な内容だったからです。Nature Plantsが創刊されることも知りませんでした。結果的に、私たちは米国のグループが投稿した1カ月後、Nature Genetics に投稿しました。するとすぐにエディターから返事がきて、今度創刊されるNature Plants へのトランスファー*を勧められました。
それで初めてNature Plants のことを知り、たいへん興味を持ちました。創刊はだいぶ先のようでしたが、トランスファーすることに決めました。Nature Plants は、Nature 関連誌の例にもれず、査読は厳しく、時間もかかりましたが、無事やりとげ、幸い掲載に至ったというわけです。投稿は遅れましたが、多数のS遺伝子の解析データを様々な角度から吟味し、しっかりとした論文に仕上げてから投稿して良かったと思っています。査読の過程で、米国のグループの研究とは全く独立した内容だと評価していただき、うれしかったですね。
―― 記念すべき創刊号の表紙に写真が掲載されましたね。
高山氏: たいへん光栄に思っています。反響も大きくて喜んでいます。Nature やNature 関連誌は採択の基準がとても公平で、内容の良さで選ぶ。そこがありがたいです。実は今、次の論文もNature Plants に投稿しようと準備しているところです。
今後は、さらにいろいろな植物を対象に、自己と非自己を見分ける仕組みを研究していきたいと思っています。また、自分の細胞と他者の細胞を見分ける動物の免疫系とも比較しながら、自己を認識する仕組みが生物全体でどのように進化してきたのかその全貌を探りたいと思っています。
聞き手 藤川良子(サイエンスライター)。
※Nature およびNature 関連誌に投稿された論文を、別のNature 関連誌に投稿し直すこと。査読を受けていた場合は、査読レポートも転送できる。
参考文献
- Takayama S., et al. Nature 413, 534-538 (2001).
- Kubo K., et al. Science 330, 796-799 (2010).
- Kubo K., et al. Nature Plants 1, Article number: 14005 (2015)
Nature Plants 掲載論文

Article: 遺伝子重複および遺伝子交換がペチュニアのS-RNase 型自家不和合性の進化を駆動する
Nature Plants 1 : 14005 doi:10.1038/nplants.2014.5 | Published online 08 January 2015
Author Profile
高山 誠司
奈良先端科学技術大学院大学 バイオサイエンス研究科 細胞間情報学研究室 教授
| 1986年 | 東京大学大学院農学研究科博士課程修了(農学博士) |
| 1986年 | 味の素株式会社中央研究所研究員 |
| 1991年 | カリフォルニア大学サンディエゴ校医学部研究派遣 |
| 1995年 | 奈良先端科学技術大学院大学 助教授 |
| 2006年 | 奈良先端科学技術大学院大学 教授(現在に至る) |

久保 健一
奈良先端科学技術大学院大学 バイオサイエンス研究科 細胞間情報学研究室 博士研究員
| 2000年 | 東北大学工学研究科博士課程後期修了(工学博士) |
| 2000年 | 農業生物資源研究所 特別研究員 |
| 2005年 | 奈良先端科学技術大学院大学 博士研究員 |
| 2011年 | 長浜バイオ大学大学院 博士研究員 |
| 2012年 | 奈良先端科学技術大学院大学 博士研究員(現在に至る) |
