日本とトルコのアブラナは、なぜ交配できない?
─自家不和合性遺伝子が作り出す生殖隔離
渡辺 正夫、髙田 美信
2017年7月号掲載
アブラナ科などの花では、雌しべに自分の花粉が付いても交配は起きない。「自家不和合性」という自己認識システムが働き、自分の花粉を拒絶するからだ。一方、他者の花粉ならば受け入れるはずである。ところが、日本系統のアブラナの雌しべはトルコ系統の花粉を拒絶するというのだ。なぜか? 今回、この発見者である東北大学の髙田美信さんと渡辺正夫さんは、拒絶の仕組みを解明し、重複した自家不和合性遺伝子が生殖隔離の仕組みに役割を果たしている可能性があることを、Nature Plants 7月号に発表した。
―― まず、自家不和合性や生殖隔離についてお聞きしたいと思います。

渡辺氏: アブラナ(Brassica rapa、図1)の花を用いて自家不和合性について研究し、30年近くになります。私の指導者で、東北大学農学部植物育種学研究室の先代教授である日向康吉博士(ひなたこうきち)(2014年逝去)が開拓された研究分野を引き継いだのです。
雌しべが、自分の花粉と自分以外の花粉を区別し、自分以外の花粉とだけ交配する。それが自家不和合性という現象です。一方、2つの集団の間で交配が起こらなくなることを生殖隔離といいます。この仕組みの解明は、科学的に興味深いだけでなく、育種学においても非常に重要です。
―― 育種学で重要とは?
渡辺氏: 自分以外の他者と交配するということは、自然界において遺伝的な多様性を生み出す仕組みとして大切な働きをしていますが、品種改良においても役立っています。品種改良では、性質の違うもの同士を交配させて新しい性質を持つ雑種を生み出したいわけですから、自分同士で交配しない性質は便利なのです。また、種苗会社が雑種の種子を得るために交配を行うときにも、便利。アブラナ科の菜の花、キャベツ、白菜、カリフラワー、カブ、ブロッコリーなどは、自家不和合性を利用して雑種の種子が採られているのです。
―― 以前に、自家不和合性を制御する遺伝子を発見されていますね。
渡辺氏: 2000年に、雌しべの先の柱頭にSRKと名付けた受容体(キナーゼ)タンパク質が発現していて、SRKが雌しべ側の自家不和合性因子であることを証明し、Natureに発表しました1。また、花粉にはその受容体に結合するリガンドであるSP11というタンパク質が発現していることも明らかにしました。
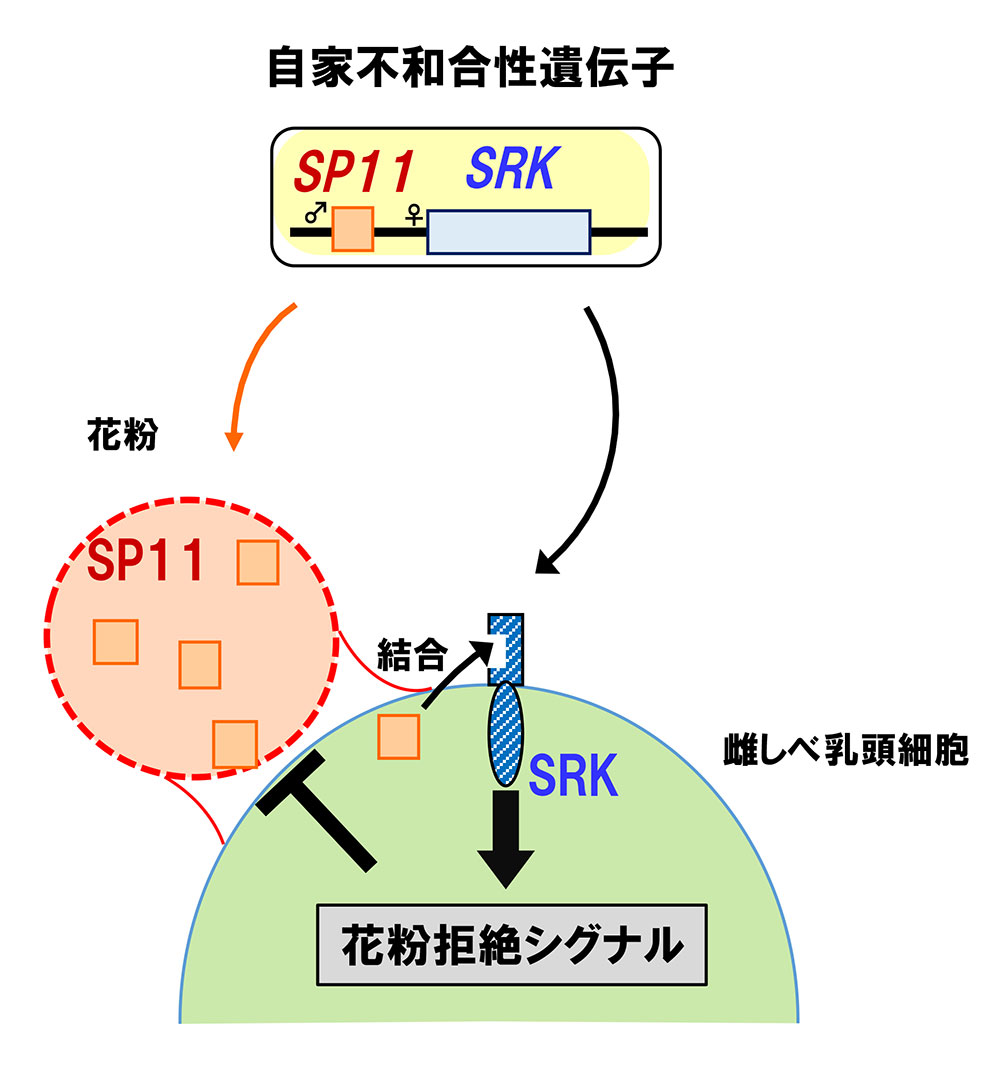
SP11遺伝子とSRK遺伝子は、染色体上ですぐ近くに並んでおり、1セットとして振る舞う。SP11遺伝子とSRK遺伝子には多数の多型(対立遺伝子)が存在する。自分のSP11遺伝子とSRK遺伝子からそれぞれ作られるリガンドと受容体が出会うと、花粉を拒絶するシグナルが発せられる。 | 拡大する
さらにその作用機序を解明し、2001年、再びNatureに発表することができました2。受容体とリガンドは、それぞれ、SRK遺伝子とSP11遺伝子とから作られます。この2つの遺伝子は染色体上で近接して位置しており、1セットのように遺伝します。これらの遺伝子には多数の多型が存在し、柱頭で自分の受容体と自分のリガンドが出会った場合、受容体とリガンドが鍵穴と鍵のようにピタッと結合してシグナルが発せられます。そのシグナルは、花粉管の伸長を阻止するように作用し、交配が起こらなくなるのです(図2)。
日本系統とトルコ系統を掛け合わせる
―― 今回、Nature Plantsに発表されたのは、その後の研究なのですね。
渡辺氏: はい。アブラナの原産地は西アジアで、私たちの温室では、原産地のトルコで採取されたアブラナも栽培しています。髙田さんがあるとき、トルコ系統の花粉と日本系統の雌しべを交配させたところ、面白いことを見つけたのです。通常でしたら、他者の花粉は受け付けるはずなのに拒絶するという現象(不和合)が起きたのです。不思議なことに、逆の組み合わせ、つまりトルコ系統の雌しべと日本系統の花粉の間では、拒絶が起きず、交配が成立しました。この現象の仕組みを解明したのが、今回の研究です。
―― これまでに知られていない現象だったというわけですか?
渡辺氏: ダーウィンの時代から、種を分ける壁は何だろうという疑問の下にいろいろな植物の交配が行われてきており、膨大な観察結果が記録・保存されています。その中に、これに似た「一側性不和合」という現象の記述がありました。ただし、一側性不和合は、異種間の現象(自家不和合性の雌しべと自家和合性の花粉との掛け合わせ)とされており、我々のアブラナの場合では、同種間の現象(自家不和合性の雌しべと花粉との掛け合わせ)という違いがありました。
―― そもそも、なぜトルコ系統と日本系統を掛け合わせたのですか?
髙田氏: 実は、偶然なのです。別の目的で実験を行っているときに、花粉が必要になりました。冬場のことで、温室で元気に咲いているのはトルコ系統のアブラナしかなく、その花粉を使ったのです。
花粉が雌しべに拒絶されたことは、受粉後に雌しべから伸びる花粉管の成長が止まることを写真撮影すれば、確認できます(図3)。しかし最初は、トルコ系統の花粉だから拒絶されたということに気づかず、研究に何年もかかってしまいました。我々の研究室では、年2回、春と秋に交配を行うというサイクルで実験を回していることもあって、実験には時間がかかってしまうのです。
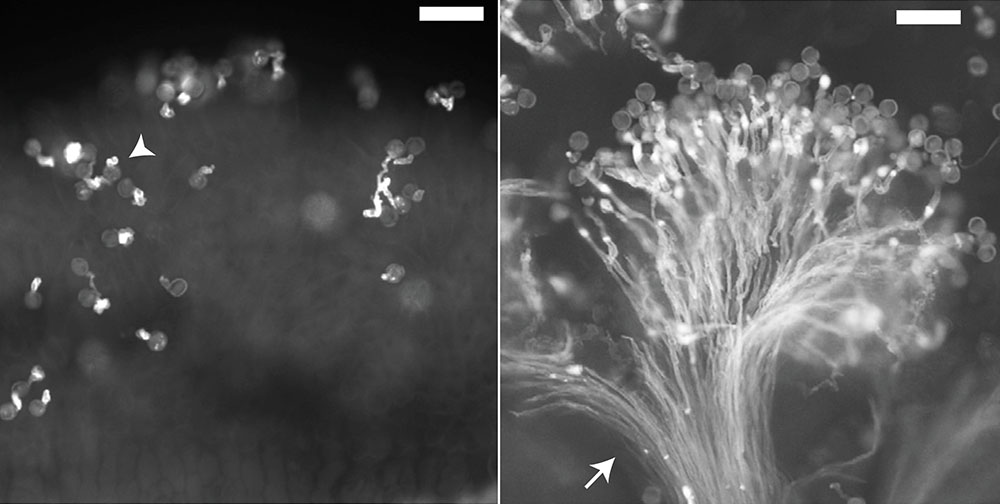
花粉管の伸長を蛍光顕微鏡で確認する(右写真、白色に見えるのが伸びた花粉管。バーは50 μm)。一側性不和合性により花粉が拒絶されると、雌しべ乳頭細胞表面で花粉発芽が起きない(花粉管が伸びない)という表現型として観察される(左写真)。 | 拡大する
―― どのように解明を進めたのですか?
髙田氏: トルコ系統を掛け合わせた実験を最初に行ったのは、2000年ごろのことでした。2005年には、拒絶(不和合)を引き起す原因が、花粉側の遺伝子1つと雌しべ側の遺伝子1つにあることを突き止めました。
次に、それらの遺伝子を具体的に特定し、花粉側をPUI遺伝子、雌しべ側をSUI遺伝子と名付けました(2013年)。偶然ですが、SUI遺伝子は、渡辺先生が助手だったときの教え子の1人であり、今回の論文のもう1人のcorresponding authorでもある、鈴木剛(すずきごう)さん(大阪教育大学教授)が1995年に発見し、報告していた遺伝子でした。PUI遺伝子は、自家不和合性のSP11遺伝子に、SUI遺伝子はSRK遺伝子に配列が似ている、ということも分かりました。アブラナのゲノム配列が2011年に解読されたのですが、それに携わった韓国のYong Pyo Limさん(忠南大学教授)からアブラナゲノムの情報を得られたことが、これらの発見に役立ちましたね。
その後、これらの遺伝子について詳細に解析し、一側性不和合が起こる仕組みを詳しく明らかにし、今回の論文としてまとめました。
―― 日本系統とトルコ系統の和合・不和合は、結局どういうことだったのですか?
髙田氏: 一側性不和合の原因遺伝子のセットは、大昔に、自家不和合性の原因遺伝子セットの遺伝子重複によって生じたと考えられます(図4)。しかしその後、遠く離れたトルコと日本で別々に変異が起こり、トルコ系統では雌しべ側の遺伝子が壊れ、日本系統では逆に花粉側の遺伝子が壊れました。したがって、それぞれの系統においては、自然界で重複遺伝子が自家不和合性に関与することはありませんでした。
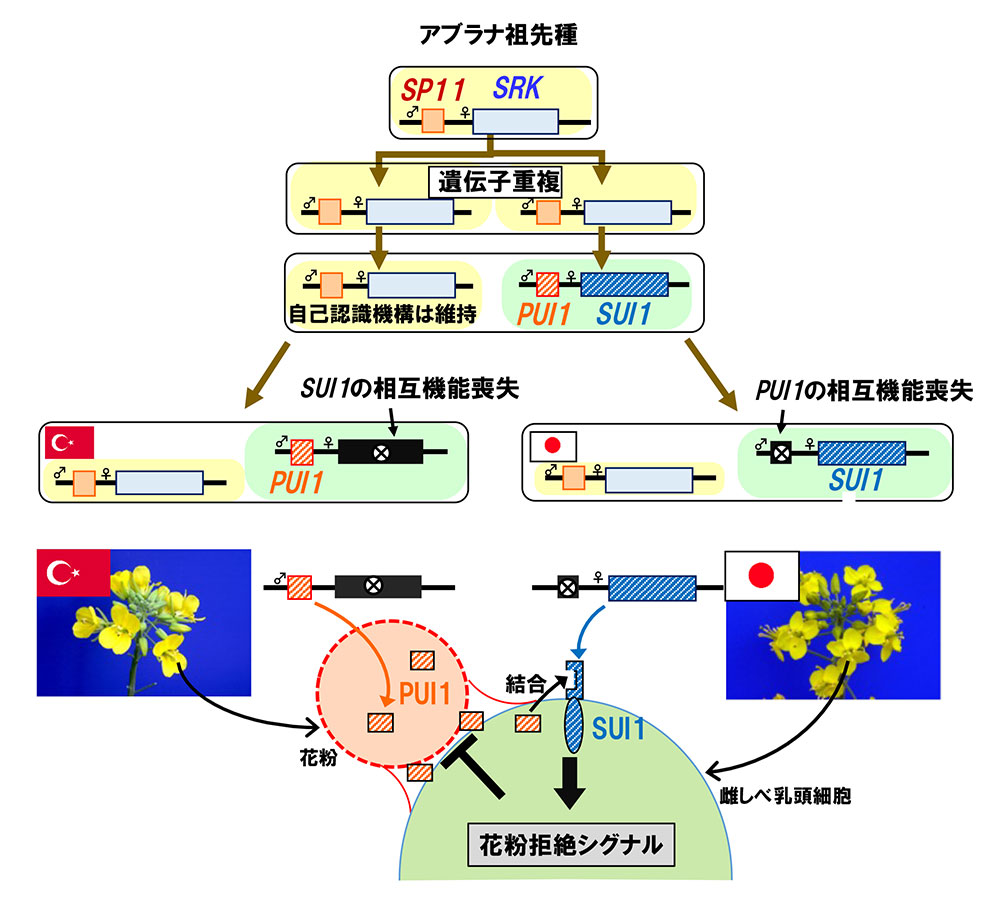
アブラナ祖先種において自家不和合性遺伝子の遺伝子重複が起こり、SUI1遺伝子とPUI1遺伝子が生み出された。その後、トルコではSUI1遺伝子が、日本系統ではPUI1遺伝子が機能喪失したと考えられる。今回、トルコ系統と日本系統を掛け合わせることにより、PUI1とSUI1が出会う組み合わせが生じ、不和合となった。 | 拡大する
ところが、我々の研究室でトルコ系統の花粉と日本系統の雌しべが出会うことになってしまい、不和合が起きる対立遺伝子のセットが生じてしまったのです(図4)。トルコ系統と日本系統の逆の組み合わせでは、それぞれの遺伝子が壊れていて、不和合が起きないというわけです。
トルコ系統と日本系統との間には、離れて生息しているうちに遺伝子重複に変異が生じ、それが原因で生殖隔離が起きつつあると考えられます。
これらの事柄は、アブラナへの遺伝子導入実験を行い、きっちり証明することができました。また、いつも共同研究を行っている高山誠司(たかやませいじ)さん(東京大学大学院教授/前奈良先端科学技術大学院大学教授)に、花粉側因子であるPUI1の合成タンパク質によるアッセイ系を教えていただき、それを使って確認実験も行いました。
―― 遺伝子重複はどんなときに起こるのでしょうか?
渡辺氏: 生物の長い進化の過程で、さまざまな偶然に基づく原因により、遺伝子重複は頻繁に起こってきたと考えられます。増えた遺伝子配列は、植物にとっては基本的に有利なことであり、そのままゲノム中に維持される傾向にあります。ただし、自家不和合性の遺伝子セットの場合には、それが重複して2セットになると、交配できる相手が狭まることになるため、おそらく生存に不利となり、重複する遺伝子の片方が変異して壊れてしまった個体が選択されだのと想像されます。
―― トルコ系統と日本系統では別々に変異が起こり、互いが交配しにくくなったのですね。これは、別々の種になる途中の過程を見ていることになるのでしょうか?
髙田氏: あくまで想像になるので、論文中には書いていませんが、その可能性はあると思います。自家不和合性遺伝子が遺伝子重複したことにより、他者(別種)として認識する潜在的な仕組みが、同種の2系統の間で形成されていたととらえることができるでしょう。つまり、新しい種が進化する途中過程かもしれないということです。
論文としてどのようにまとめたらいいか
―― 進化とも関連してくるということですか?
渡辺氏: 実験データは、文句のつけようのないほどきれいに証明ができていたので、あとは、論文をどうまとめるかとなったときに、進化の話題にどれほど深く踏み込むかについては迷いました。しかし結局は、論じなかったのです。髙田さんが鈴木さんと具体的に検討を行ったのですが、種の進化に関する「ドブジャンスキー・ミュラーのモデル」にも触れませんでした。このモデルのような、流行のキーワードを論文に入れたいという思いはあったのですが……。
髙田氏: このモデルは、「遺伝子重複後に2つの集団に別々の変異が起こり、それにより2集団の間での交配で生まれた子の遺伝子に不適合があると、これら集団間に交配後隔離が成立する」というものです。ゲノム進化に詳しい清水健太郎(しみずけんたろう)さん(スイス・チューリッヒ大学准教授)に相談したところ、今回の現象はこのモデルと比較して論じることが可能であろうとのことでした。しかし、ジャーナルのレフェリーと議論になったときに、それに応じることができるかと考えると、自分の専門を超えると感じました。それで、論文では論じないことにしたのです。
―― Nature Plants を投稿先に決めたきっかけは?
渡辺氏: 我々としては、Nature 系のジャーナルのどれかには、いけそうだと感じていました。まずはNature に投稿したのですが、結果はリジェクトでした。
次の投稿先の選択肢の1つとして、Nature Communications がありました。オープンアクセスジャーナルですから、誰もが読めると思ったからです。しかし結局、植物生殖遺伝学という自分たちの専門に最も近いことから、髙田さんがNature Plants を選びました。
髙田氏: Nature Plants のエディターやレフェリーからは、びっくりするほど好意的なコメントが返ってきて(笑)、掲載がすんなりと決まりました。もっとも、「進化のことをもっと書いてください」というレフェリーからのコメントを見たときには、ちょっと後悔の念がよぎりました。進化について論じていれば、Nature でも、最初の扉が開いたかもしれないと思えて。
―― 渡辺さんは、これまでNature に何度も論文が掲載されていますが、論文執筆で心がけていらっしゃることは?
渡辺氏: どういう順番で内容を執筆していくかと考えるときに、Nature は万人が読むジャーナルであるということを意識しますね。専門外の人が興味を持ってくれるためには、科学の背景や歴史に関する言葉から書きはじめるのがよいというのが、高校生などにも授業をした経験からいえることです。例えば、「メンデルは……」「ダーウィンは……」というようにね。
―― 高山さんとはいつも共同研究を行っていらっしゃるとのことでしたが……。
渡辺氏: 高山さんの師匠である奈良先端科学技術大学院大学の磯貝彰(いそがいあきら)名誉教授と私の師匠の日向先生の時代から、共同研究を行ってきた間柄です。磯貝研究室は生化学・生物有機化学が専門で、微量の活性物質を抽出するのが得意な、いわゆる「物取り屋」さん。磯貝先生の出身研究室(東大農学部)の先達である田村三郎(たむらさぶろう)博士(2015年逝去)は、大量のタケノコから植物ホルモンのジベレリンを抽出したことで有名な方です。我々は、自家不和合性に関する因子などの抽出をお願いしてきました。
専門性の高い分野は専門家にお願いするというのが、日向先生の時代からの我々の方針です。今回も、タンパク質のアッセイ系は高山さん、進化については清水さん、遺伝子マーカーについては諏訪部圭太(すわべけいた)さん(三重大学准教授)、アブラナゲノムについてはLimさんというような共同研究をすることで、この研究が成功したと思っています。もちろん、大量の実験をこなし、最も貢献したのは髙田さんですが。髙田さんは、アブラナの花粉症にならないのです。
―― アブラナで花粉症が起きるのですか?
渡辺氏: 私の知っている研究者の中で、アブラナの花粉症にならなかったのはたった2人。髙田さんと日向先生だけです。
髙田氏: 春には、温室の中が花でいっぱいになります。そこで作業をすると、体まで黄色くなるくらいです。このため、たいていの人は何年かすると花粉症になってしまうので、マスクをするようにしています。でも、私は全く平気なのです。
アブラナの実験の大変さは、花粉症だけではありません。遺伝子導入実験(形質転換)が容易ではないという点もあります。今回の研究では、アブラナの芽生えから切り出した胚軸の数が10万個をはるかに上回りました。今回の研究に15年以上を要したことには、こうした理由も関係しています。
―― 今後の研究の展開は?
髙田氏: 進化の研究に関しては、他の方が進めてくれるでしょう。私は、トルコと日本のそれぞれの系統で、変異して壊れた部分の遺伝子配列を元に戻すことができないだろうかと考えています。そうすれば、自己不和合性遺伝子のセットを2つ手に入れることになります。遺伝子セットを2つ持つことは、自然界では不適合でしたが、野菜などの栽培においては、自家不和合性を安定させるために役に立つ可能性があるのです。
渡辺氏: さらに付け加えれば、栽培種と野生種との生殖隔離の方法として応用できるといいですね。
―― ありがとうございました。
参考文献
- Takasaki, T. et al. The S receptor kinase determines self-incompatibility in Brassica stigma. Nature 403, 913–916 (2000).
- Takayama, S. et al. Direct ligand-receptor complex interaction controls Brassica self-incompatibility. Nature 413, 534–538 (2001).
インタビューを終えて
渡辺先生の話は止まらない。書き留めたインタビューのメモが何ページにもわたった。「渡辺先生は、エネルギッシュですね。若い時とちっとも変わりません」と髙田さんも言う。研究に加えて、高校生などへの理科教育といったアウトリーチ活動も大切にされ、あちこち飛び回るそうだ。他に大切にされていることはありますかと尋ねたところ、即座に黄ばんだ本を取り出された。何かと思ってみると、戦前の植物学の書籍。そして、「このような古い文献にも丁寧に当たること」とおっしゃった。そのような書籍・文献の中には、当時の技術の限界から、事実の解明には至らなかったものの、驚くほど詳細な観察結果や洞察力に富んだ私見が見られるとのこと。それが、今でも研究のヒントやアイデアとなるらしい。
聞き手は、藤川良子(サイエンスライター)。
Nature Plants 掲載論文

Letters: 重複した花粉・雌ずい認識座位がBrassica rapaの種内一側性不和合性を制御する
Nature Plants 3 : 17096 doi:10.1038/nplants.2017.96 | Published online 26 June 2017
Author Profile
渡辺 正夫(わたなべ まさお)
東北大学大学院生命科学研究科植物生殖遺伝分野 教授
| 1990年 | 東北大学大学院農学研究科博士課程前期修了 |
| 1991年 | 東北大学農学部 助手 |
| 1994年 | 博士(農学)取得 |
| 1997年 | 岩手大学農学部 助教授 |
| 2005年 | 東北大学大学院生命科学研究科植物生殖遺伝分野 教授(現職) |

髙田 美信(たかだ よしのぶ)
東北大学大学院生命科学研究科植物生殖遺伝分野 技術職員
| 2005年 | 岩手大学大学院連合農学研究科 博士課程修了(博士(農学)) |
| 2005年 | 大阪教育大学 研究員 |
| 2005年 | 東北大学大学院生命科学研究科 研究員 |
| 2007年 | 東北大学大学院生命科学研究科植物生殖遺伝分野 技術職員(現職) |
