細胞内共生が起源? 葉緑体が植物の成長を制御する未知の仕組み
増田 真二
2015年12月号掲載
葉緑体は、約15億年前に光合成細菌が原始真核生物内に共生することで誕生したとされる。このほど、共生の際に細菌から持ち込まれたRSHという遺伝子が、葉緑体や植物体の大きさを制御する機能を持つことを、東京工業大学バイオ研究基盤支援総合センターの増田真二准教授らが明らかにした。
―― 葉緑体が植物体の成長を制御するという斬新な内容です。
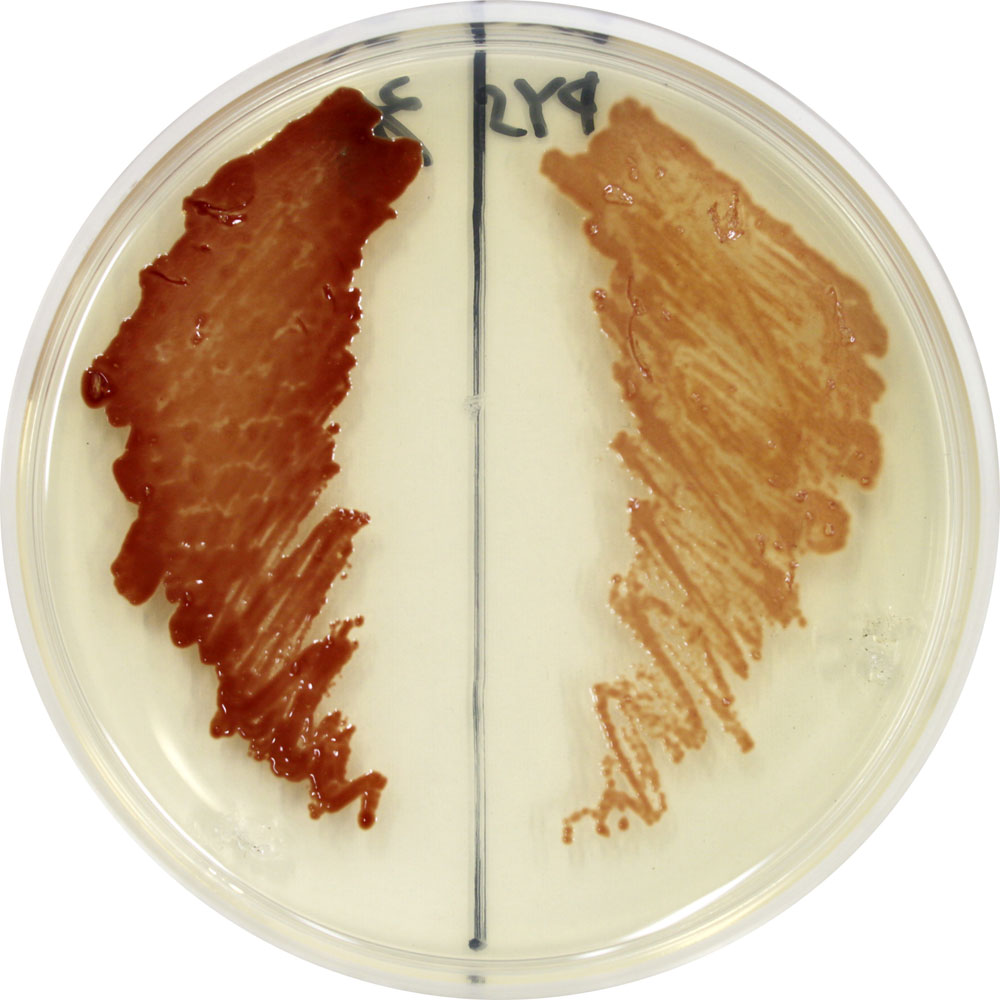
色の薄い変異体はppGpp合成/分解酵素を欠損しており、緊縮応答不全となっている。 | 拡大する
増田氏: はじめから植物の成長にフォーカスしていたわけではありません。もとは、紅色細菌などを対象に、光エネルギーや光情報の利用メカニズムを研究していました。紅色細菌は有酸素下では酸素呼吸を、無酸素環境では光合成を行う細菌で、葉緑素のほかカロテノイドを多く持っているために赤い色をしています。ところがあるとき、色の薄い変異体を見つけました(図1)。不思議に思って調べてみると、遺伝子発現や代謝酵素の活性を低く抑える仕組み(緊縮応答)が不全状態にあり、緊縮応答が呼吸と光合成のバランス制御に重要な役割を果たしていることがわかりました。
この研究を論文にした後、2005年に東工大の助手として、植物を扱っていた高宮建一郎先生の研究室に赴任しました。その少し前に、国際的なゲノムプロジェクトによって、配列上はシロイヌナズナにも紅色細菌と同じ緊縮応答のための遺伝子があることがわかり、それなら植物での機能も調べてみようと考えました。あれから10年。今回、緊縮応答が葉緑体で起こること、その結果として植物体の成長や大きさが制御されることなどが明らかになったのです。
―― 緊縮応答とはどのような現象なのでしょうか?
増田氏: 環境変動に対応するための仕組みで、シアノバクテリアから大腸菌まで細菌界に広くみられます。細菌は、環境が悪化し、アミノ酸欠乏などの低栄養状態に陥ると、ATPとGTPから「グアノシン4リン酸(ppGpp)」を作り出します。このppGppがシグナル分子として機能すると、細胞増殖にかかわる遺伝子発現が抑えられ、逆に、核酸やアミノ酸代謝に関連する遺伝子発現が上昇します。効率よく代謝して生き延びる生存戦略で、栄養状態がよくなるとppGppは分解されて元に戻ります。これまでの研究から、ppGppの合成を触媒する酵素遺伝子として、relAとspotの2つが知られています。このうちSpoTはppGppの合成だけでなく、分解も担うことがわかっています。先の研究で私は、紅色細菌のSpoTが光合成と呼吸の調節にも重要であることを突き止めました1。
2000年代に入ると、シロイヌナズナのゲノムにもrelAやspotと似た遺伝子があることがわかり、RelA/SpoT Homologs の頭文字をとってRSH遺伝子と名付けられました。興味深いのはその起源です。葉緑体は、光合成細菌であるシアノバクテリアが「原始的な植物細胞」に取り込まれ、細胞内共生したことで誕生したとされます。このときrelAやspotも葉緑体ゲノムに存在していたはずです。その後、葉緑体ゲノムから核ゲノムに移動しRSHとなりましたが、タンパク質自体は今でも葉緑体で機能しています。私には、植物が偶然手にした緊縮応答機能を、自分に都合のよいように飼いならしたように思えます。
―― 今回はシロイヌナズナのRSHについて、機能解析をされたわけですね。
増田氏: はい、そのとおりです。シロイヌナズナにはRSH1、RSH2、RSH3、CRSH (Ca2+-activated RSH)の4種のRSH遺伝子があります。私たちはそのうちの2種(RSH2とRSH3)について、欠損変異体と過剰発現体を作り、葉緑体などの表現型を詳しく調べてみました。その結果、欠損変異体では特に異常がみられなかったのですが、過剰発現体では次のように多くの異常がみられました。
まず、葉緑体についてです。数は変化しませんでしたが、大きさが小さくなりました。一方、葉緑体中のppGppは、野生型の4〜5倍も多く蓄積していました。次に植物体についてです。なぜか成長が加速し、通常の約1.5倍の大きさに達しました。ただし、色は野生型よりも薄くなり、黄緑色でした。最後に耐貧栄養性ですが、ある程度成長すると、窒素栄養が少ない状態でも生き延びられるようになりました。これらの結果から、RSHが葉緑体と植物の成長や大きさを制御する機能を発揮していると結論づけ、論文をNature Plants に投稿しました。

緊縮応答を過剰に引き起こす組換え植物体は、通常条件下では、葉緑体のサイズは減少するが、個体は大きく育つ。この組換え体を窒素欠乏条件下にさらすと、緑色を保ち、光合成を継続した。 | 拡大する
―― 生き延びるための仕組みで大きくなるとは不思議な気がします。
増田氏: Nature Plants 編集部も、その点に興味を抱かれたようです。私にも確固たる理由はわかりませんが、RSHは単細胞の緑藻類などにも保存されていることから、本来は葉緑体を制御するために機能するもので、大きくなったというシロイヌナズナの表現型は二次的にもたらされたに過ぎないのかもしれません。
RSH過剰発現体では、植物体内のメタボローム解析(全代謝産物を網羅的に解析する手法)もしました。こちらは予想どおりで、アミノ酸をはじめとする多くの代謝産物が減っていることがわかりました。減った分を成長に回したのだと思います。種子も作らせてみたところ、内部のアミノ酸含有量などが低いためか、発芽率が低下することがわかりました。次世代のことまで考慮できない状態になっているのかもしれません。
―― 論文はすんなり受理されたのでしょうか?
増田氏: 解析のタイミングについて、「より早い成長段階のものを加えるように」といった指示はありましたが、大方すんなりいきました。論文掲載に当たって News & Views にも取り上げられ、英国の研究者から「葉緑体は植物に大きな利益をもたらすが、その維持には膨大なコストをかける必要があり、RSHはこの利益とコストのバランスに重要なのではないか」とコメントをいただきました。光合成などの葉緑体機能を人工的に強化することは、植物バイオマスの増大に必ずしも結びつくとは限らない、という点で注目してくださったのでしょう。
―― 一連の研究で、どこが評価されたとお考えでしょうか?
増田氏: 緊縮応答の遺伝子発現や代謝制御が、高等植物では興味深い表現型に寄与することを突き止めた点があげられますが、質量分析の手法を駆使することで「高精度にppGppを検出する系」を確立できた点も評価されたと考えています。これまでは、うまく定量する手法がありませんでしたので。
この研究の応用として、植物の大きさの制御技術や植物工場開発なども考えられ、地球環境や食糧問題の観点で興味を引いた面もあるかもしれません。すでにいくつか特許出願をしていますが、応用は共同研究で行い、個人としては基礎研究を続けていきたいと考えています。
―― 今後の目標は何でしょうか?
増田氏: つい最近、フランスのチームが「RSH1〜3の三重変異体を作成したが、ppGpp量に大きな異常はみられなかった」と報告しました。この三重変異体は私たちも解析していたところで、先を越されたかたちです。4つ目のCRSHが最も重要だろうと考えられますが、この遺伝子を欠損した変異体は存在しません。そこで現在は、最近開発が進んだゲノム編集技術を使ってCRSH欠損変異体の作製に着手しています。同じことをフランスのチームも考えていると思うので、機能解析まで含めて急がなくてはと思っています。ゆっくり進めたかったのですが、そうもいかないですね。あまり先のことを考えるタイプではないので、次の目標は今の研究が一段落したら考えたいと思っています。
ただし、ppGppの局在については、調べられたら興味深いと考えています。RSHを細胞質で特異的に発現させると植物体が小さくなることがわかった2ので、ppGppが細胞質にもあると推測しているのですが、直接的に局在部位を調べる方法がないのです。葉緑体以外の部位で、どのように機能するのか解析したいと思うのですが。
―― 最後に、若手研究者へのアドバイスを一言お願いします。
増田氏: 最近は、同じようなデータを使って書いても、文章表現を工夫しないと有名ジャーナルには論文が受理されにくいと感じています。一方で、十分なデータを持ちながら、なかなか論文化を進めない若手研究者が多いように思います。自戒を込めて言うと、なるべく多くの論文を書いて文章力をみがいてほしいと考えます。はじめから高いレベルを狙わずに、少し気楽に書く練習を積んではどうかと思います。
聞き手 西村尚子(サイエンスライター)。
参考文献
- Masuda, S. & Bauer, C.E., J. Bacteriol. (2004) 186: 235-239.
- Ihara,Y. & Masuda, S., Plant Signal. Behav. (2016) e1132966.
Nature Plants 掲載論文

Article: 植物の生育およびストレス応答に認められるプラスチドの緊縮応答の影響
Impact of the plastidial stringent response in plant growth and stress responses
Nature Plants 1 : 15167 doi:10.1038/nplants.2015.167 | Published online 9 November 2015
Author Profile
増田 真二
東京工業大学バイオ研究基盤支援総合センター基盤部門 准教授
| 2000年 | 東京都立大学理学研究科生物学教室博士課程修了 博士(理学)取得 |
| 2000年 | 日本学術振興会特別研究員(PD) |
| 2000年 | インディアナ大学客員研究員 |
| 2003年 | 理化学研究所基礎科学特別研究員 |
| 2004年 | 東京工業大学生命理工学研究科助手 |
| 2008年 | 東京工業大学バイオ研究基盤支援総合センター基盤部門准教授(現職) |
