食虫植物の進化がゲノム解読から明らかに
長谷部 光泰、福島 健児
2017年3月号掲載
植物なのに、虫を捕らえ、食べる食虫植物。この不思議な生き物は、いったいどのように進化してきたのだろうか。このほど、自然科学研究機構基礎生物学研究所の長谷部光泰さんと米国コロラド大学の福島健児さんらは、食虫植物フクロユキノシタのゲノム配列を明らかにし、さらに捕らえた虫を分解する消化酵素の進化について解明して、Nature Ecology & Evolution 3月号に発表した。食虫植物の進化の謎解きに挑むお二人に聞いた。
―― 食虫植物とは、どのような植物ですか。

長谷部氏: 虫を「食べる」ことにより、窒素やリンなどの栄養素を吸収しているのが、食虫植物です。「食べる」というのは、その葉を用いて、虫などの小動物を誘引、捕獲、消化、吸収することです。通常の植物はこうした栄養素を根から吸収するのですが、食虫植物は虫などからも吸収できるので、貧栄養の土壌でも生育が可能です。
―― 虫を捕らえるための器官は、葉なのですね。
長谷部氏: そうです。捕虫葉と呼ばれますが、普通の葉と同じように茎の先端部分に作られる葉原基から発生します。一口に捕虫葉といっても虫の捕らえ方はさまざまで、モウセンゴケの粘り付け式や、ウツボカズラの落とし穴式、ハエトリソウの挟み込み式など、いろいろあって面白いですよ(図1)。私も福島君も、もともと食虫植物の面白さに引かれていて、大好きです。そういう人は多く、趣味で食虫植物を育てている愛好家が世界中にたくさんいます。

ウツボカズラやヘイシソウ(サラセニアともいう)は落とし穴式、モウセンゴケは毛の先端から粘液を分泌する粘り付け式、ハエトリソウは挟み込み式だ。ハエトリソウは、一定の時間内にトゲに2回物が触れると、葉が閉じる。物が触るという記憶を持つことができる。 | 拡大する
―― 捕虫葉の形は実に多彩ですね。
長谷部氏: 捕虫葉の独特の形状がどのように作られるかについては、今回の研究に先立って解析を進めてきました。通常の平面葉の場合には、形作りの概要が分かりつつあります。葉のへりにある表と裏の境目部分の細胞が分裂しやすくなっているのです。そして、細胞の分裂面が葉の表面に対して垂直になるように分裂することによって、葉が横に広がっていきます。
では、捕虫葉の袋は、どのようにして袋の形となったのか。従来考えられていた仮説は、葉の裏側が表側に食い込むことにより、丸くなり、袋状になるのではないかというものでした。しかし、そうではないことを福島君が発見したのです。
福島氏: ムラサキヘイシソウ(ヘイシソウはサラセニアともいう)という食虫植物を用いて調べたのですが、葉の裏側を特徴づける遺伝子が、表側では発現していないことに気づきました。つまり、この仮説が成り立たないということです。そこで、葉の細胞の分裂面がどのような方向を向いているのか、徹底的に調べました。すると、葉の付け根にある細胞の分裂面の方向に、他と違っているところがあるのを見つけたのです。そこの細胞分裂を、同じ基礎生物学研究所の藤田浩德さんたちの協力でコンピューターシミュレーションしたところ、平らな葉にへこみができることが分かりました(図2)。
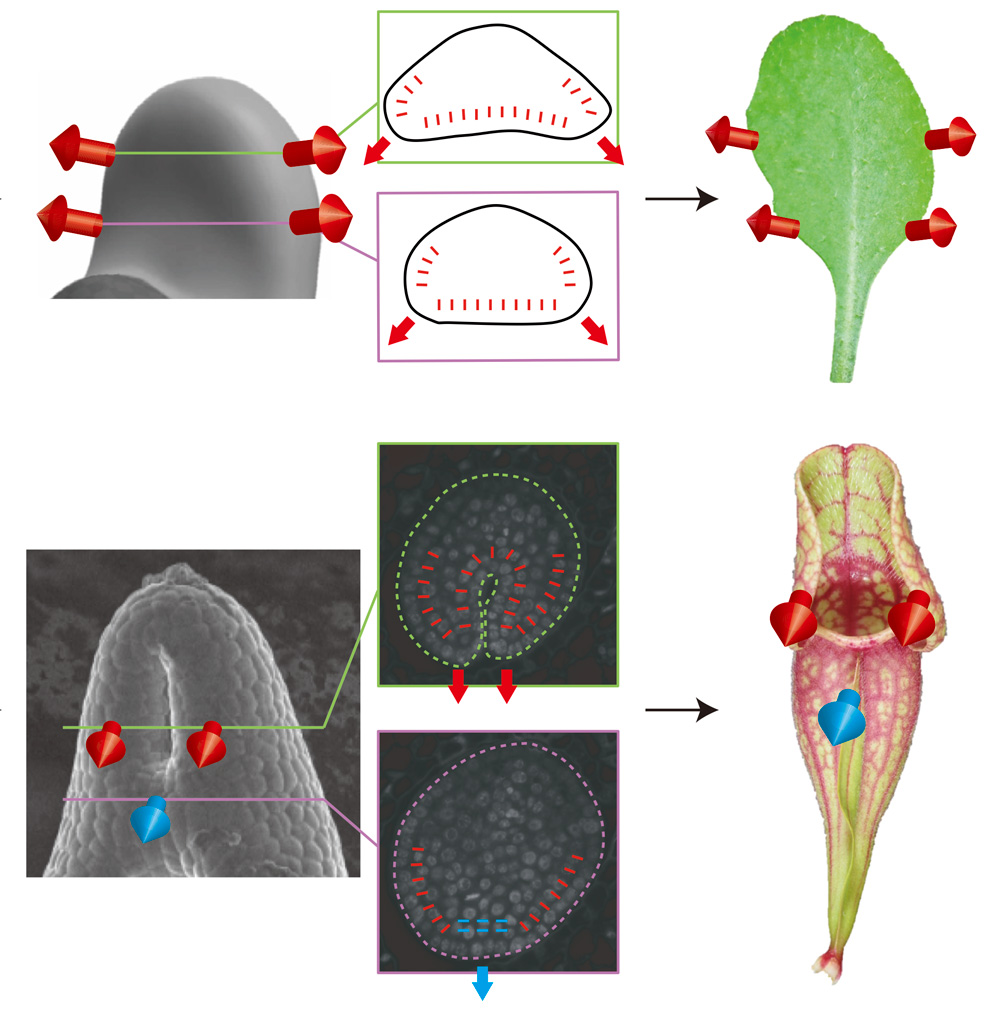
分裂面の方向の違いを赤と青の線で示した。平面葉では、葉の表面に対して分裂面が垂直(赤色線)だが、平行の分裂面が生じて(青色線)、両者が組み合わさるとへこみができ始める。 | 拡大する
長谷部氏: 福島君ががんばって、小さな葉の断面切片を何千枚も作り、観察したのですよ。論文にまとめ、2015年にNature Communications に掲載することができました。
しかし、分子レベルでさらに研究を進めていくためには、食虫植物のゲノム解読を行って、遺伝子の情報などを解析に利用できるようにすることが重要だと痛感しました。
フクロユキノシタを研究材料に選ぶ
―― 今回は、フクロユキノシタを研究されたのですね。

1つの株に捕虫葉と平面葉の両方がつく。スケールは1cm。 | 拡大する
福島氏: はい。オーストラリアに生育するフクロユキノシタCephalotus follicularis(図3)を研究材料に用いて、ゲノム配列を解読し、さらに遺伝子を調べることにより食虫性の進化について明らかにしました。研究材料選びは慎重に行いました。すでに、オオバナイトタヌキモという食虫植物のゲノムが解読されていましたが1、我々はフクロユキノシタの方がより食虫性進化の研究に適していると考えました。フクロユキノシタは、1つの株に捕虫葉と平面葉(通常の葉)の両方がつくタイプの食虫植物であり、遺伝子の機能などを捕虫葉と平面葉で比較でき、都合がよいのです。ただ、普通の栽培条件下では捕虫葉と平面葉を作り分けることはできません。しかし今回、いろいろ工夫して作り分けに成功し、研究用に使えるようになりました。
―― 捕虫葉と平面葉を作り分けるとは?
長谷部氏: 捕虫葉も平面葉も、茎の先端にある葉原基から発生するのですが、発生の初期段階では、顕微鏡で見ても両者を識別できません。それでは、例えば、捕虫葉の発生時に発現する遺伝子を調べたいといっても、難しいですよね。
福島氏: 葉の作り分けがうまくいかなかった頃、食虫植物愛好家の知恵を借りようとしたことがありました。栽培に詳しい方が多いので、何か秘けつをつかんでいるのではないかと考えたのです。ところが、私が尋ねた限りでは、そのような方法はないとのこと。がっかりしましたが、それなら自分で見つけようと、栄養、日照、温度など、さまざまな生育条件を設定して実験を試みました。そして最終的に、白色光の連続照射下で、栽培温度によりほぼ100%作り分けられることを発見しました。15℃では平面葉のみが、25℃では捕虫葉のみができたのです(図4)。基礎生物学研究所のインキュベーターをたくさん借りて実験したかいがありました。

15℃だと平面葉に、25℃だと捕虫葉になる。 | 拡大する
長谷部氏: 葉の作り分け方が分かるという保証がない中で、よく見つけられたものだと、驚きましたよ。後から考えれば、進化的にもっともだと思うのですが。高温の環境の方が、虫が多いですからね。温度が発生を切り替えるスイッチになるというのは、野外の植物である程度使われている仕組みなのかもしれません。
ゲノム解読から消化酵素の進化解明へ
―― そして、フクロユキノシタのゲノムを解読されたのですね。
福島氏: はい。フクロユキノシタのような袋状の捕虫葉は、その中に消化液がためられています。(捕虫葉を傾けながら)この透明な液体がそうです(図5)。消化液には消化酵素が何種類か含まれていて、虫が中に落ちると分解されます。

今回、消化液中のタンパク質と捕虫葉で発現している遺伝子の両方を調べることで消化酵素の遺伝子を突き止め、その配列を明らかにしました。
さらに、この遺伝子の配列を詳細に見ていくと、面白いことが確認できました。普通の植物(非食虫植物)にもウイルスや細菌を殺す病害抵抗性酵素が備わっているのですが、食虫植物の消化酵素というのは、大昔に、こうした病害抵抗性酵素が変化して生じた可能性が大きいと分かったのです。これまで、一部の酵素については他の研究者からも推測されていましたが、今回、ゲノム全体を解析することで、そのほぼ全容を明らかにできました。
―― フクロユキノシタ以外の食虫植物の消化酵素ではどうなのでしょうか。
福島氏: 世界には数百種の食虫植物が知られていますが、それらは被子植物の5つの異なる系統群、イネ目、カタバミ目、ナデシコ目、ツツジ目、シソ目にみられます。つまり、全く別々に進化を遂げてきた異なる系統の植物から、同じような形質が独立して進化しているわけで、このような現象を収斂(しゅうれん)進化といいます。
今回は、3つの異なる系統に属する4種(3目4種)の食虫植物で、消化酵素を比較してみました。フクロユキノシタ(カタバミ目フクロユキノシタ科)の他に、ヒョウタンウツボカズラ(ナデシコ目ウツボカズラ科)、アデレーモウセンゴケ(ナデシコ目モウセンゴケ科)、ムラサキヘイシソウ(ツツジ目ヘイシソウ科)です。これらの食虫植物が持つ消化液中の消化酵素を精製して比較したところ、互いによく似た酵素が使われていることが分かりました(図6)。
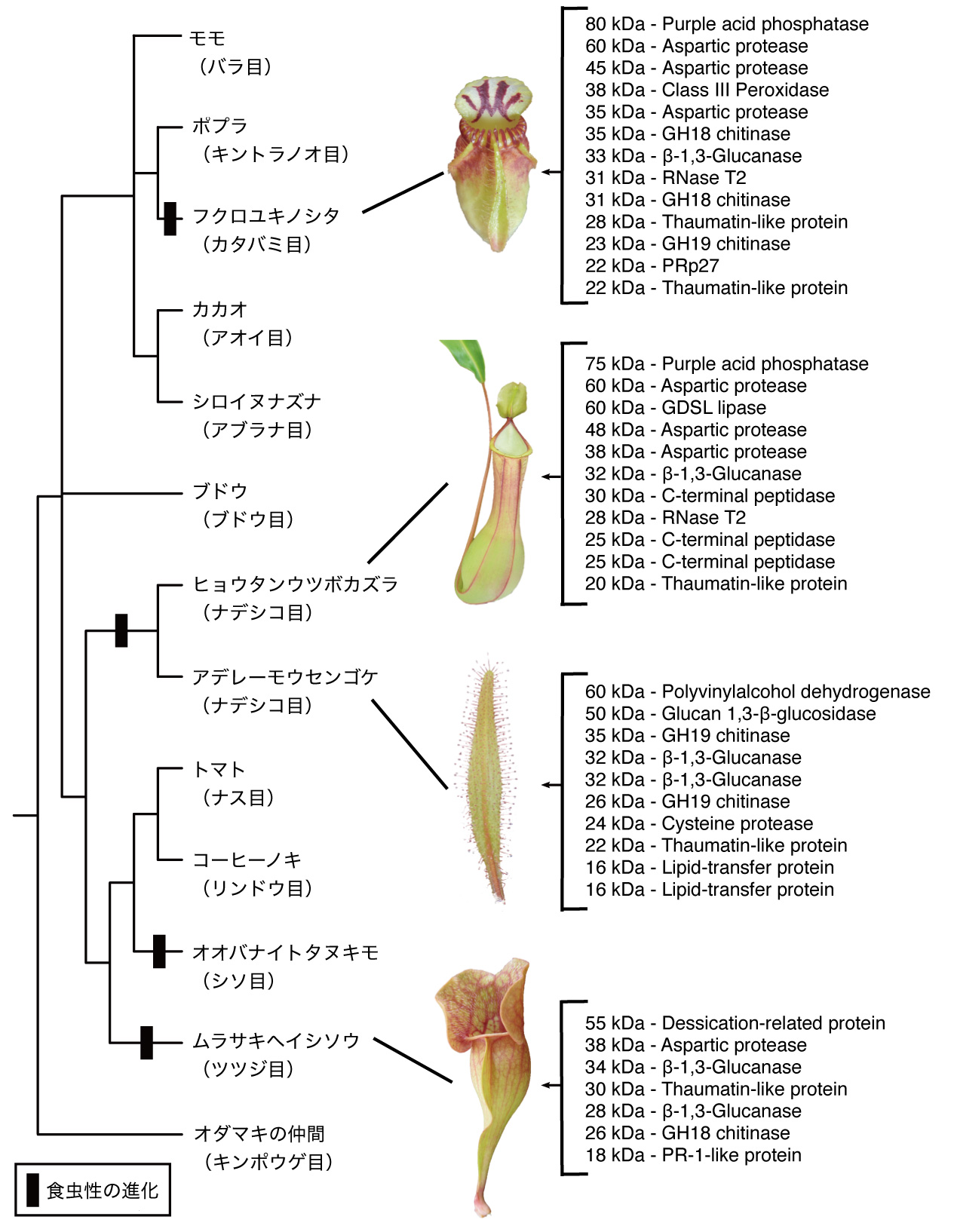
フクロユキノシタ(カタバミ目フクロユキノシタ科)、ヒョウタンウツボカズラ(ナデシコ目ウツボカズラ科)、アデレーモウセンゴケ(ナデシコ目モウセンゴケ科)、ムラサキヘイシソウ(ツツジ目ヘイシソウ科)。系統樹の右側は、含まれていた酵素の種類。 | 拡大する
その1つ、キチン分解酵素(キチナーゼ)を例にみてみましょう(図7)。どの植物のゲノムにも、この分解酵素に相当する遺伝子が10個程度含まれています。異なる系統で、それぞれ独立して進化してきた食虫植物であるならば、約10個の遺伝子から異なるものが選ばれてもよさそうですが、実際には、フクロユキノシタとヒョウタンウツボカズラで同じ1つの遺伝子が選ばれて、消化液中の酵素に用いられているのです。すなわち、系統が異なっても、食虫性を進化させていく過程で、同一の病害抵抗性に関わると推定される分解酵素を利用してきたということです。酵素の遺伝子レベル(分子レベル)でも収斂進化が起こっていたことに驚きました。食虫植物に進化するには、限られた道筋しかなかったのだともいえるのではないでしょうか。
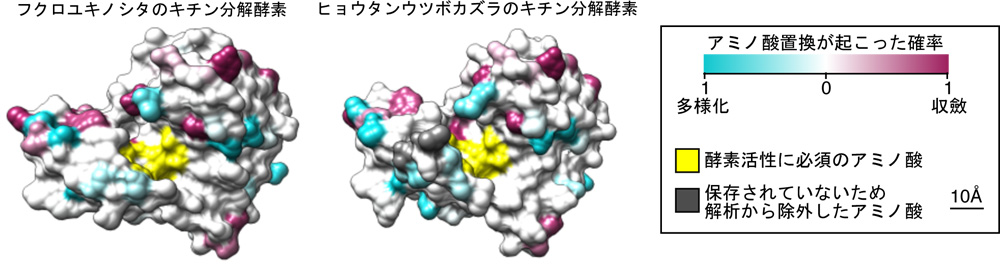
黄色はこの酵素の活性に必須のアミノ酸であり、両者は同じ。青色はアミノ酸置換により、別のアミノ酸に変化した(多様化)。紫色もアミノ酸置換が起こったが、同じアミノ酸に変化した(収斂進化)。 | 拡大する
―― 進化の道筋が限られていたとは、どういうことなのですか。
長谷部氏: 消化酵素を捕虫葉の消化液中で機能させるには、いろいろな制約があります。消化液中にはタンパク質を分解する酵素も含まれているので、それにより消化酵素自体が分解されてはいけないし、また、常温で活性が保たれなければなりません。そういった制約がある中で機能させるには、特定の病害抵抗性酵素が適しており、またこれを利用するしかなかったのではないかと考えています。
―― 消化酵素のほかに、ゲノム解読から明らかになった食虫性形質はありますか。
福島氏: 捕虫葉と平面葉が作られるときに、それぞれどのような遺伝子が発現しているかを調べた結果、消化酵素以外にも、食虫性に関わると考えられる遺伝子の候補が多数見つかりました。例えば、葉の形に関する遺伝子の候補は130ほどあります。
―― 今回の研究はNature Ecology & Evolution の3月号に掲載されましたね。
福島氏: Nature Ecology & Evolution が創刊されることは、昨年からSNSなどで話題になっていました。このようなジャーナルが創刊されて、とても喜んでいます。エボデボ(発生進化学)の論文も対象になるので、うれしいですね。
長谷部氏: Nature Ecology & Evolution のエディターは、研究の内容を非常によく理解されていて、驚きました。コミュニケーションもスムーズに運びました。また論文掲載後は、いろいろな国のニュースで取り上げていただき、海外からの問い合わせも多数きています。
―― 今後はどのように研究を展開されるのですか。
長谷部氏: 捕虫葉には、今回解析した点以外にも、虫を捕獲するための工夫がいろいろ見られます。それがどのように進化してきたのかについても、明らかにしていきたいですね。例えば、フクロユキノシタの捕虫葉のふたには透明な部分があり、そこが明るく見えるので虫がおびき寄せられます。さらに、袋の口の部分は滑りやすく、おびき寄せられた虫が袋に落ちるような構造になっています。面白い仕組みがたくさんあるのですよ。
こうした食虫性に関わる仕組みはさまざまに進化してきましたが、完成形に至るまでの途中段階ではどうなっているのか。それも、解明したいですね。生物のある特徴(食虫性など)は、進化の途中段階では、一時的に生存に不利な状態が起きていると考えられますが、それをどう乗り越えているのかは、まだ解明できていない進化学の課題なのです。例えば、葉が袋の形になれたとしても、消化や吸収の仕組みを獲得するまでは、その葉は光合成の効率という点から生存に不利です。複雑な形質がどのように進化していったのか、食虫性の進化を解明していくことで、明らかにしていければと思います。今回、フクロユキノシタのゲノムデータが得られたことから、その情報を利用して、研究が大きく発展していくことでしょう。捕虫葉に関する福島君の研究は、その後、留学生のGergo Palfalvi君が引き継いでくれていますが、フクロユキノシタのゲノムデータを用いて遺伝子レベルの解析が進められます。
福島氏: 私は長谷部研で学位を取得した後、コロラド大学で収斂進化の研究をさらに専門的に進めています。植物よりも動物の方が研究に利用できるゲノムデータが豊富に存在するので、今は動物を研究材料としています。収斂進化の情報学的解析技術を開発し、それをまた植物のフィールドにフィードバックしたいと願っています。
―― ありがとうございました。
参考文献
- Ibarra-Laclette et al., Nature, 498, 94–98 (2013)
インタビューを終えて
福島先生がコロラド大学から一時帰国したタイミングに合わせて、長谷部研究室でお話を伺った。長谷部研には植物の巨大乾燥標本がたくさん飾られており、目を奪われる。例えば、エンドウ豆を大きくしたようなさや入りの種子。長さは1m近い。高さ30cmほどの松ぼっくりもある。長谷部先生の面白い解説が次から次へと始まる。「この松ぼっくりはオーストラリア産。たくさんついているカサの1つ1つに種が入っているんだけど、このカサってなんだと思う? 遺伝子を調べたらはっきりわかったんだけど、枝が進化して縮まったものなんですよ」、といった具合。
長谷部先生が目指すのは、面白い講義、面白い研究だ。「基礎科学って、すぐには役に立たない分、面白くなくちゃ。できるだけ大勢の人に面白いと思ってもらえる研究をやりたい。真理の発見は、みんなも面白いはずだもの」。
聞き手は、藤川良子(サイエンスライター)。
Nature Ecology & Evolution 掲載論文

Article: 食虫植物フクロユキノシタのゲノムから明らかになった、食虫性に関連する遺伝的変化 OPEN
Genome of the pitcher plant Cephalotus reveals genetic changes associated with carnivory
Nature Ecology & Evolution 1 : 0059 doi:10.1038/s41559-016-0059 | Published online 6 February 2017
Author Profile
長谷部 光泰(はせべ みつやす)
大学共同利用機関法人自然科学研究機構 基礎生物学研究所生物進化研究部門 教授
| 1991年 | 東京大学大学院理学系研究科博士課程植物学専攻 3年次中途退学 |
| 1991年 | 東京大学大学院理学系研究科附属植物園岩槻邦男研究室 助手 |
| 1992年 | 博士(理学)取得 |
| 1993年 | パデュー大学(米国)日本学術振興会海外特別研究員 |
| 1996年 | 岡崎国立研究機構(当時)基礎生物学研究所種分化機構第2研究部門 助教授 |
| 2000年 | 大学共同利用機関法人自然科学研究機構 基礎生物学研究所生物進化研究部門 教授(現職) |
| 2000年 | 総合研究大学院大学 教授(兼任) |

福島 健児(ふくしま けんじ)
コロラド大学デンバー校アンシュッツ医療キャンパス(米国)研究員
| 2015年 | 総合研究大学院大学生命科学研究科基礎生物学専攻5年一貫制博士課程(基礎生物学研究所生物進化研究部門)修了(博士(理学)) |
| 2015年 | コロラド大学デンバー校アンシュッツ医療キャンパス(米国)日本学術振興会海外特別研究員 |
| 2017年 | コロラド大学デンバー校アンシュッツ医療キャンパス 研究員(現職) |
