記憶T細胞形成のカギを握るのは、脂質代謝
中山 俊憲、遠藤 裕介
2019年2月号掲載
麻疹やおたふく風邪などは、一度かかると、再び発症することはほとんどない。これは、免疫細胞が病原体のタンパク質を記憶していて、再感染の際に速やかに応答しているからだ。このような免疫記憶の中心となる記憶T細胞の産生に脂肪酸代謝が関わっていることを、千葉大学大学院医学研究院教授で研究院長の中山俊憲さんとかずさDNA研究所オミックス医科学研究室室長の遠藤裕介さんらの研究グループが発見し、新創刊のNature Metabolism 2月号に発表した1。
―― まずは、免疫記憶についてお聞きします。
中山氏: 体内に侵入した病原微生物が増殖を始めると、感染症を発症します。これに対し、その微生物のタンパク質(抗原)に特異的な抗体が増加し、感染源を排除します。こうして感染症は治ります。その後再感染すると、まるで抗原を覚えているかのように迅速な免疫応答が起こり、発症しなかったり、かかっても症状が軽かったりします。これが免疫記憶という現象で、それを利用したのがワクチンです。ワクチンの始まりは、220年ほど前のジェンナーによる種痘です。当時、牛痘(牛の天然痘)にかかった牛飼いは、天然痘に感染しないことが経験的に分かっていました。そこで、子どもの腕に傷を付けて牛痘の膿を塗り込んだところ、天然痘にかからなかったのです。このようにワクチンは、病原微生物の抗原を体内に入れて免疫記憶として覚えさせるのです。
―― 免疫記憶の中枢を担っているのが、記憶T細胞というわけですね。
中山氏: はい。図1を見てください。抗原に出会ったことのないCD4+T細胞をナイーブCD4+T細胞といいます(以下、CD4+を略す)。ナイーブT細胞は、病原微生物の感染によって抗原提示細胞に示された抗原を認識して活性化し、Th1やTh2、Th17などの生体防御の最前線で働くエフェクターT細胞になり、それぞれ異なる役割を担います。いうなれば、ナイーブT細胞は国家試験に受かったばかりの新米医師、エフェクターT細胞は臨床研修を終え、内科や皮膚科、外科という専門性を身につけた専門医というわけです。これらの細胞のほとんどは死滅しますが、ごく一部は記憶T細胞として、感染源が排除された後も体内に数カ月から年単位で維持されます。そして再感染すると、初回よりも迅速に活性化されて、病原体を速やかに排除します。このため、記憶T細胞をより長くより多く維持できれば、長期間効果が持続するよいワクチンになります。ただ、その所在やメカニズムはまだよく分かっておらず、我々もさまざまな切り口で研究しています。その一環で、病原性(pathogenic)の記憶T細胞がいることも分かってきました。
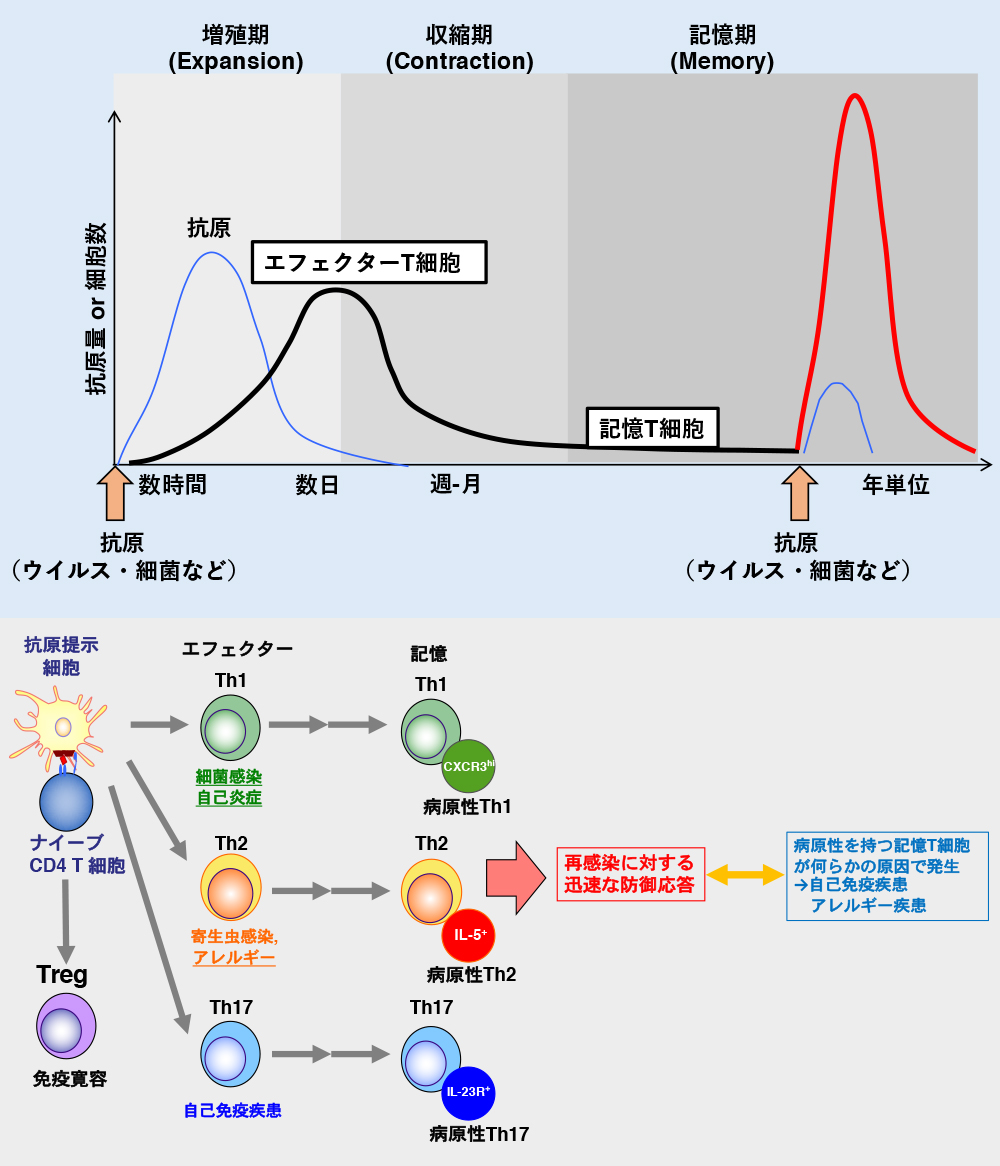
免疫記憶は、ナイーブT細胞が外来抗原を認識し、一過性に増殖・エフェクター機能を獲得することから始まる。記憶T細胞は免疫記憶の司令塔であるにもかかわらず、どのような特徴を示す細胞が、いかなるメカニズムにより生体内で長期間生存するのかはいまだ明らかになっていなかった。 | 拡大する
―― 病原性記憶T細胞とは?
中山氏: 喘息やリウマチなどの難治性のアレルギー疾患や自己免疫疾患では、記憶細胞をなくしてしまえば、症状が良くなることが分かっています。我々は最近の研究で、線維化を起こす難治性の喘息で有害な病原性記憶T細胞とその分泌物を同定しました2。
―― 今回は、代謝変化という切り口での研究ですね。
遠藤氏: 休止期のナイーブT細胞がエフェクターT細胞に分化すると、劇的に増殖してサイズも大きくなるので、栄養素を細胞外から取り込んだり細胞自身で作り出したりしなければなりません。そして、休止期の記憶T細胞になると、再び小さくなり増殖しなくなります。これだけ性質が変わるのに代謝が変わらないわけがないと思ったのです。
―― 初めから脂質代謝を見据えていたのですか?
遠藤氏: いいえ。いろいろな代謝について、数十種類の阻害剤を用いて実験しました。阻害剤の濃度検討から始めて、とにかく大変で、脂質代謝にたどり着くまで3年くらいかかりました。実は、この研究を始めた頃は、免疫と代謝の関係はあまり言及されておらず、中山教授がこの実験を認めてくれたことが大きいです。そして、結果が出るまで根気強く待ってくれました。
中山氏: 広い心と長い目です(笑)。
―― 脂質代謝関連酵素の中で、ACC1(アセチルCoAカルボキシラーゼ)を取り上げましたが。
遠藤氏: 遺伝子発現のプロファイリングを行ったところ、脂質代謝関連酵素の遺伝子の発現が、ナイーブT細胞や記憶T細胞で低くて、エフェクターT細胞で高いことが分かりました。ACC1をコードする遺伝子(マウスではAcaca)もその1つで、脂肪酸生合成で重要な律速酵素です。ACC1の阻害やノックアウトしたエフェクターT細胞から効率よく記憶T細胞が産生されました。野生型の約5倍増え、かつ抗原の再刺激でも増えたのです。
―― どういうメカニズムで記憶T細胞様の細胞になったのでしょう?
遠藤氏: エフェクターT細胞では、解糖系の活性が高く、TCA回路や脂肪酸のβ酸化の活性は相対的に低い状態です。ナイーブT細胞や記憶T細胞では、その逆です。こうした代謝変化をメタボリックリプログラミングといいます。ACC1阻害やノックアウトしたエフェクターT細胞では、TCA回路やβ酸化が上昇し、記憶T細胞に近い状態になっていました。また、これまでの研究からACC1を阻害して脂肪酸合成を抑えるとβ酸化が亢進される結果が得られています。このことから、ACC1阻害やノックアウトしたエフェクターT細胞は、ACC1の阻害によりβ酸化が促進され、その代謝産物を使うTCA回路が効率よく回って記憶T細胞のような代謝プロファイルになっていると考えられます。こうして記憶T細胞のように振る舞えるようになったエフェクター細胞は、より生存できるようになり、メタボリックリプログラミングがスムーズに行われるようになって、効率よく記憶T細胞になっているのでしょう。
―― 今回、記憶T前駆細胞の同定という大きな成果が得られましたね。
遠藤氏: はい。ほとんどのエフェクターT細胞は脂質合成が盛んですが、一部は脂質合成が低くなっていてそれが記憶T前駆細胞ではないかと想定しました。そこで、単一細胞解析を行ったところ、エフェクターT細胞には、脂質合成酵素の発現が高い細胞と低い細胞があることが分かりました(図2)。このデータが出た時は、うれしくて飛び上がりそうなくらいでした。
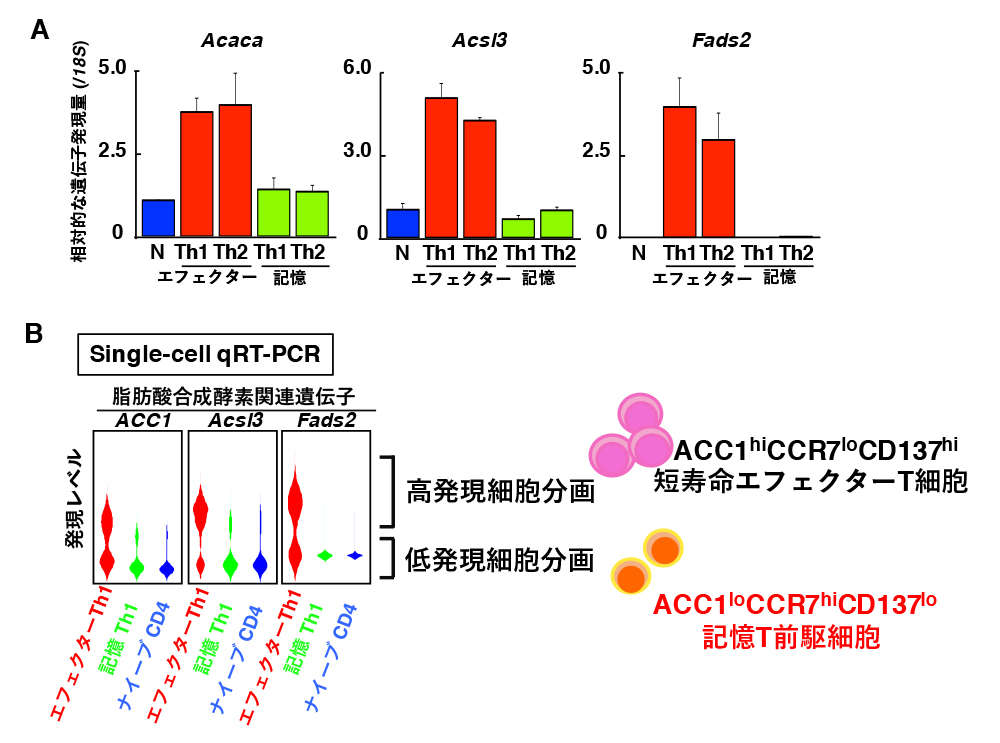
A. 通常のqRT-PCRによる脂肪酸合成酵素の発現量。エフェクターT細胞でこれらの酵素の発現が上昇し、記憶T細胞で低下していた。
B. 単一細胞qRT-PCRによる脂肪酸合成酵素の発現量。エフェクターT細胞の中に、発現の低い細胞集団がいることが分かった。ACC1を指標にした統計解析により、ACC1低発現細胞群ではCCR7の発現が高く、CD137の発現が低いことが示された。
注:qRT-PCR法
RT-PCRは、逆転写(RNAを鋳型としてDNAを合成すること)により合成されたDNA(cDNA)を用いてPCRを行う方法である。qRT-PCRは蛍光プローブを用いて、PCR増副産物をリアルタイムでモニタリングし、より定量的に解析する手法である。本手法により、タンパク質をコードする遺伝子を選択的に増幅したり、少量しか存在しないmRNAを増幅したりすることが可能となった。最近の研究では、非常に小さな同集団の細胞であっても実際には不均質性が存在することが明らかになっており、単一細胞qRT-PCRを行えば、単一細胞における遺伝子・RNA発現量が分かる。 | 拡大する
次に、脂質合成の低い細胞をうまく分けられないかと思い、脂質合成に相関または逆相関する遺伝子を調べました。こうした細胞では、脂質合成に相関する遺伝子の発現が低くなり、逆相関する遺伝子の発現は高くなると考えたからです。選んだのは、細胞表面分子のCCR7とCD137をそれぞれコードする、Ccr7(逆相関)とTnfrsf9(相関)です。CCR7の発現が高くCD137の発現が低い(CCR7hiCD137lo)細胞をセルソーターで分画して、記憶T細胞形成能を調べたところ、予想通り、効率よく記憶T細胞になりました。さらにCCR7hiCD137lo細胞は、脂質合成酵素の発現が低く、記憶T細胞に近い代謝プロファイルであることも分かりました。
以上のことから、エフェクター細胞の中には、脂質合成が低い記憶T前駆細胞が存在すると決定づけました。
中山氏: 他の代謝経路ではなく、脂質合成が低いことが記憶T前駆細胞の特徴である事実が重要です。
遠藤氏: 長年やってきたので、本当に喜びひとしおでした。
中山氏: この知的エクスタシーが忘れられないんですよ。それを求めて次の研究を進め行く。一種の依存症です。
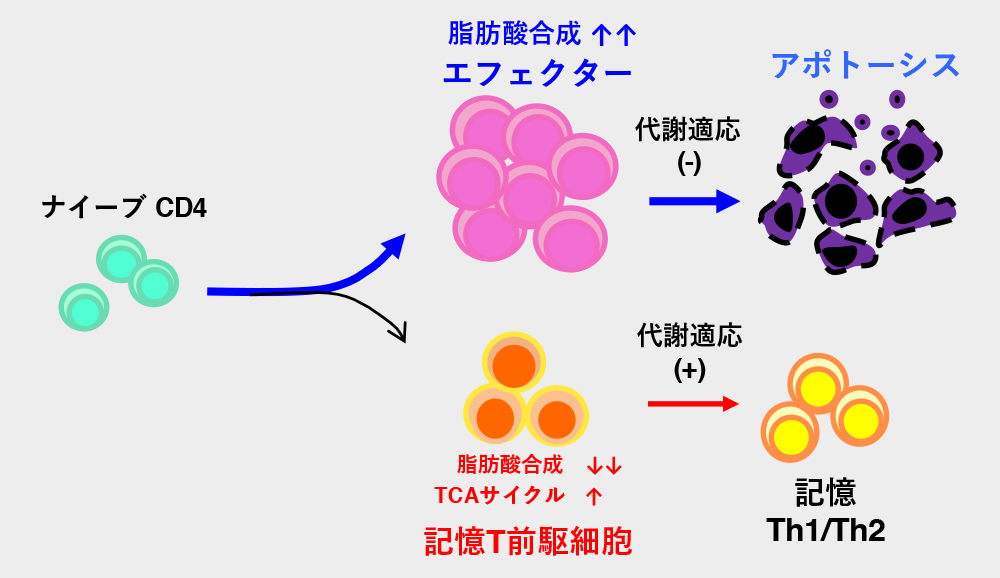
エフェクターT細胞には脂肪酸合成が盛んな短寿命エフェクターT細胞と、脂肪酸合成酵素の発現が低く、記憶T細胞へ効率よく分化することのできる記憶T前駆細胞が存在する。今後、脂肪酸代謝の調節で記憶T細胞を人為的にコントロールして優良な記憶T細胞だけを増やすことができれば、強力で長期間効果のあるワクチンの開発が可能である。一方、生体にとって有害となる病原性の記憶T細胞を減らせれば、慢性炎症の治療も期待される。 | 拡大する
―― ところで、免疫学に興味を持ったきっかけは?
中山氏: 私は、医学部生の頃から実験をしたいと思っていて、実は医師の研修もしませんでした。だから、採血よりネズミの尻尾への注射の方が得意なんです。私のMDはMedical Doctorではなく、Mouse Doctorです(笑)。免疫学を選んだのは、当時、1980年代初頭にはよく分からないことが多い免疫学が面白そうだと思ったからです。
遠藤氏: 私は化学科の出身ですが、生命現象の研究に関わりたいと思っていました。特にT細胞やB細胞は同じ細胞なのに表現型が劇的に変わることを知り、非常に興味を持ちました。その頃、中山教授がT細胞の機能変化とエピジェネティクスの研究を始めていて、すごい面白いことをやっているなと思い、修士に応募しました。実際、研究室に入ったら本当にすごく面白くて、現在に至っています(笑)。
―― 今後はどのようなことを研究したいですか?
中山氏: 1つは、病原性記憶T細胞がどうやって通常の記憶T細胞から再分化するのかということです。病原性記憶T細胞にも前駆細胞があるのかなど、疑問は尽きません。もう1つは、以前から行っているもので、分化したTh1細胞やTh2細胞がどうやってクロマチンを制御して分裂後も同じサイトカインを出す細胞として維持されるのかという研究です。どちらのテーマも基本的には何も分かっていないので、興味は尽きません。
遠藤氏: 各免疫細胞の「顔」となる脂質と脂質代謝酵素の全てを見つけ、何が免疫細胞の個性を規定しているのかを知りたいです。免疫の分野では脂質はあまり研究されていなく、全然分かっていません。中山教授の言葉を借りれば、「代謝で免疫を制御する」ことを最終目標にしています。
―― 最後に研究を目指している若い人へのメッセージをお願いします。
中山氏: まず大学生には、大学は自由な時間があるので、得意なところを伸ばすチャレンジを勧めます。医療も研究もチームで行います。チームでは自分に得意なところがなければ他人に頼られない、自分がいい仕事をしないと共同でもいい仕事ができないのです。
個人的には、臨床医より研究者の方が面白いと思っています。分からないことへの素朴な疑問の答えを求めることが、研究のモチベーションになっています。ちゃんと実験をやっても、10回のうち1回くらいしか予想通りの結果にならないことがほとんどです。でも、その成功の喜びが、次の努力につながっていく。そしてこの知的エクスタシーは、もう1回あると増強されます(笑)。
遠藤氏: 研究で1番楽しいことは、世界中でいまだ誰も知らないことに取り組めることです。科学に興味があるのなら、まずはトライです。そして、早い時期にポジティブデータを体験できれば、きっと研究にはまります。
―― ありがとうございました。
参考文献
- Endo,Y. et al. ACC1 determines memory potential of individual CD4+ T cells by regulating de novo fatty. Nature Metabolism 1, 261-275 (2019)
- Yamamoto, T. et al. DUSP10 constrains innate IL-33-mediated cytokine production in ST2 hi memory-type pathogenic Th2 cells. Nature Communications 9, 4231 (2018)
インタビューを終えて
インタビューはとにかく興味深く、全て記しきれないのが残念です。終始和やかな雰囲気で行われましたが、それも中山先生のお人柄によると思いました。遠藤先生も良い先生に巡り会えたことが大きいとコメントしていました。少年のような探求心で研究を続けておられる中山先生、そして遠藤先生。ともにサイエンスの醍醐味は、分からないことへの答えを見つけることだと言われていました。まさに、その通りだと感じたインタビューでした。
聞き手は、田中明美(サイエンスライター)。
Nature Metabolism 掲載論文

Article: ACC1は脂肪酸合成経路を介して記憶CD4+T細胞への運命決定を制御する
Nature Metabolism 1 261–275 doi:10.1038/s42255-018-0025-4 | Published online 14 January 2019
Author Profile
中山 俊憲(なかやま としのり)
千葉大学大学院医学研究院教授、同研究院長・医学部長
千葉大学副学長
カリフォルニア大学医学部サンディエゴ校 兼任教授
| 1984年 | 山口大学医学部卒業 |
| 1988年 | 東京大学大学院医学系研究科修了(医学博士) |
| 1988年 | 米国国立がん研究所 客員研究員 |
| 1991年 | 東京大学医学部免疫学教室 助手 |
| 1995年 | 東京理科大学生命科学研究所 助教授 |
| 1998年 | 千葉大学大学院医学研究院 助教授 |
| 2001年 | 同 教授 |
| 2014年 | 千葉大学副学長(現職) |
| 2015年 | 千葉大学大学院医学研究院長・医学部長(現職) |
| 2018年 | カリフォルニア大学サンディエゴ校 医学部兼任教授(現職) |
主な受賞歴:第3回日本免疫学会賞(2000年)、第14回アボットジャパン・アレルギー学術奨励賞(2004年)など。
免疫記憶に関する生物学と病原性Th2細胞の分化の分子機構を解明することが現在の最重要課題と思って、研究している。

遠藤 裕介(えんどう ゆうすけ)
公益財団法人かずさDNA研究所先端研究開発部オミックス医科学研究室 室長
千葉大学大学院医学研究院オミクス治療学 特任准教授(兼任)
| 2005年 | 慶應義塾大学理工学部応用化学科卒業 |
| 2007年 | 千葉大学医学研究科医科学修士課程修了 |
| 2011年 | 千葉大学医学薬学府博士課程修了(博士(医学)) |
| 2011年 | 千葉大学医学研究院免疫発生学教室 特任研究員 |
| 2012年 | 千葉大学医学研究院免疫発生学教室 特任助教 |
| 2013年 | 千葉大学医学研究院免疫発生学教室 特任講師 |
| 2018年 | 公益財団法人かずさDNA研究所先端研究開発部オミックス医科学研究室 室長(現職) 千葉大学大学院医学研究院オミクス治療学 特任准教授(兼任、現職) |
主な受賞歴:第64回日本アレルギー学会学術集会Featured Poster賞、第44回日本免疫学会ワークショップベストプレゼンテーション賞、第11回日本免疫学会研究奨励賞など。
明らかになっていることとなっていないことを正確に整理し、“Conceptual Advance”へ挑戦することを楽しみ、そして“代謝で免疫を制御する”ことを最終目標にしている。
