大規模な比較トランスクリプトーム解析で、ミトコンドリアの縮退機序を探る!
橋本 哲男、神川 龍馬、稲垣 祐司
2017年4月号掲載
生物の進化系統樹上には、一度獲得したミトコンドリアの機能を二次的に縮退させた真核微生物が散見される。残された構造体は「ミトコンドリア関連オルガネラ(MROs)」と総称されるが、その大きさ、失った機能、残された機能等は実に多様で、謎も多い。今回、筑波大学の橋本哲男さんらは5カ国からなる国際チームを組み、嫌気性の真核単細胞生物の一群、メタモナス類に属する18種を対象に大規模な比較トランスクリプトーム解析を行い、ミトコンドリアを縮退させた道筋の一端を明らかにした。
―― ミトコンドリア関連オルガネラとはどういうものでしょうか?

橋本氏: 真核生物にみられるミトコンドリアは、酸素を利用してエネルギー源のATPを産生する酸素呼吸(好気呼吸)をはじめ、アミノ酸代謝、鉄・硫黄クラスター合成、酸化ストレス応答などの機能を担う細胞内小器官(オルガネラ)です(図1)。太古の地球には酸素がなく、生物は嫌気環境のもとで誕生しました。やがて光合成をして酸素を産生する微生物が現れ、好気環境へと変わってきました。すると今度は、「好気的なエネルギー産生能を獲得した原核生物(真正細菌)」が現れ、それが真核生物の祖先となる微生物内に共生してミトコンドリアになったと考えられています。ところが、嫌気環境や寄生環境に適応している真核微生物の中には、獲得したはずのミトコンドリアを縮退させたものが存在しています。ただし、すべてを捨て去っているわけではなく、生息環境に応じて一部の構造や機能を残しており、それらを総称して「ミトコンドリア関連オルガネラ(Mitochondrion Related Organelles(MROs))」と呼んでいます。
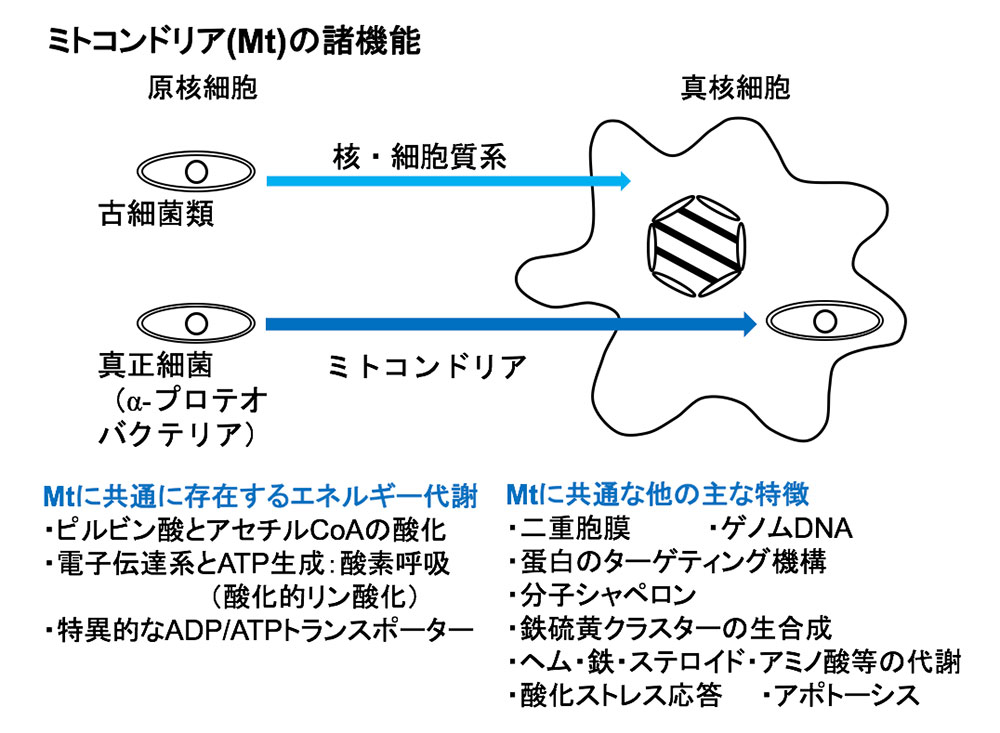
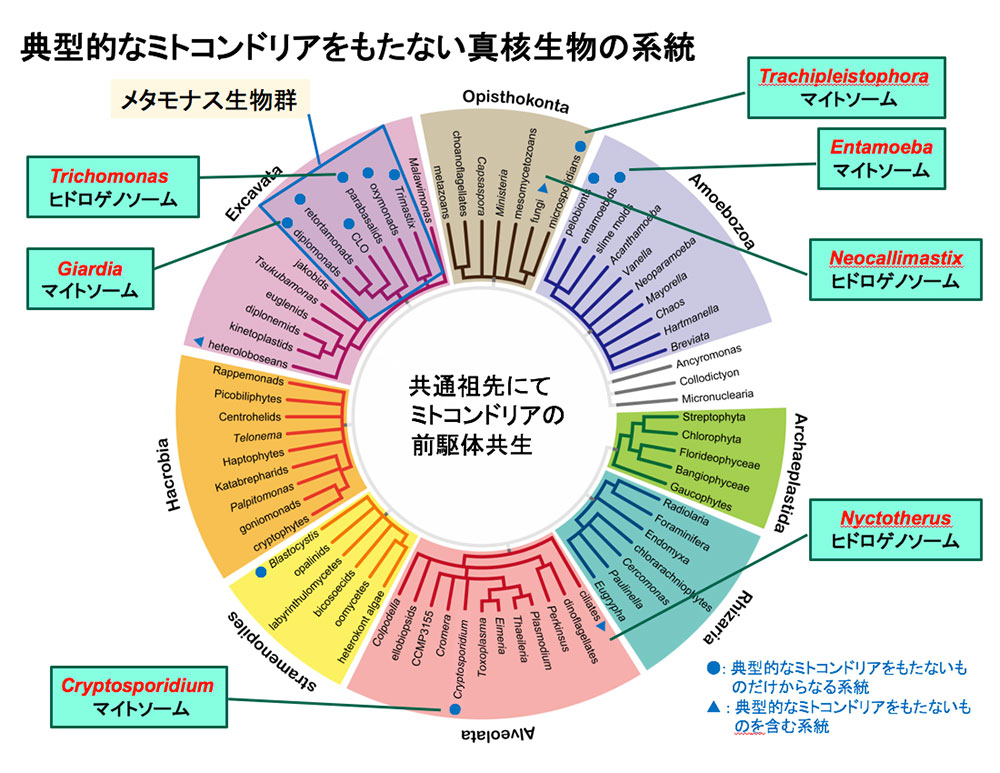
実は、MROについてはよく分かっておらず、謎が多く残されています。ごく最近まで、十分な実験が行われず、また解析技術がなかったからです。例えば、トリコモナス(Trichomonas)やジアルディア(Giardia;ランブル鞭毛虫)という真核微生物は、ミトコンドリアをもたないと考えられてきました。そしてこうした真核微生物は、完全なミトコンドリアをもたないことを根拠の1つとして、「原始的」とされてきたのです。そんな中、2000年初頭、トリコモナスの水素産生オルガネラであるヒドロゲノソームがミトコンドリアの痕跡をとどめていることや、ジアルディアもマイトソームと呼ばれる「機能がほとんど残っていないMRO」をもつことが報告されました。加えて、次世代シーケンサーの登場により高速かつ高精度にDNAやRNAの配列が読めるようになり、系統学的位置をより正確に推定できるようになった結果、原始的なのではなく、「一度獲得したミトコンドリアの機能を捨てた状態にある」ことが分かってきたのです(図2)。
―― これまでのご研究について簡単にご紹介ください。
橋本氏: 私たちの研究室では、トリコモナスやジアルディアなどの嫌気環境を好む真核微生物を対象に研究を進めてきました。トリコモナスは膣内に生息する性感染症病原体、ジアルディアは腸管に生息する旅行者下痢症の病原体として知られています。これまでに、どちらも核ゲノム内に「ミトコンドリアで機能する遺伝子」をもつことなどを突き止めてきました。例えば、「タンパク質の立体構造化に関与するシャペロン遺伝子」や「鉄・硫黄クラスター合成遺伝子」などです。さらに、これらの遺伝子の分子系統解析を行うと、トリコモナスもジアルディアも「ミトコンドリアをもつ一般的な真核生物グループ」の中に入ってしまうこと、つまり、通常のミトコンドリアで機能しているものと同一起源の遺伝子であることが明らかになりした。
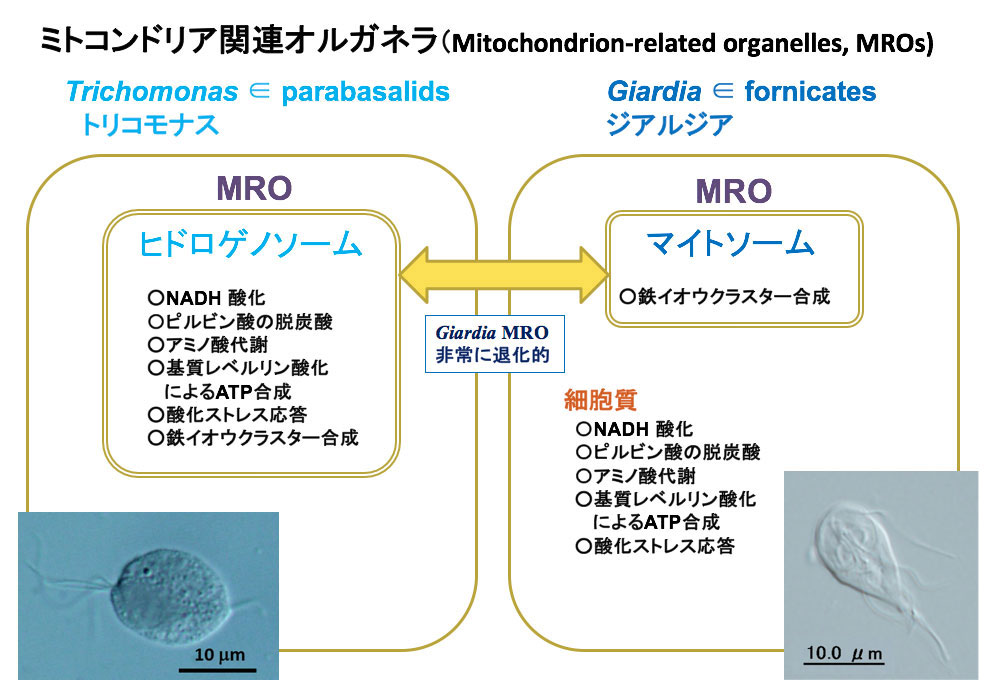
同じ頃、トリコモナスのヒドロゲノソーム(直径約0.5μm)がMROであることや、ジアルディアも鉄イオンクラスター合成系のタンパク質が局在する極小サイズのオルガネラ(直径約0.2μm)を持ち、それもMROだとする報告がなされました。トリコモナスはヒドロゲノソーム中で水素を利用してATPを産生していますが、ジアルディアのMROはATP産生能を失い、細胞質において解糖系の一部として酢酸からATPを作り出しています(図3)。
これらの知見は、トリコモナスよりもジアルディアの方がミトコンドリアの縮退が顕著だということを示しています。そこで今回は、系統的に両者の間に位置する微生物を対象に、転写産物の大規模な解析(トランスクリプトーム解析)を行い、どのように縮退が進んだのかを推定しようと考えたのです。
―― 具体的にどのようなことをされたのでしょう?
橋本氏: トリコモナスとジアルディアは共に、メタモナス類という鞭毛(べんもう)を持つ単細胞微生物の一群に含まれます。そこで、系統関係において両者の間に位置する11種を合わせた計18種のメタモナス類(フォルニカータ生物群12種、パラアバサリア生物群3種、プレアクソスティラ生物群3種)を対象に、大型計算機を用いた大規模なトランスクリプトーム比較解析を行いました。18種は、土壌や海・河川などの水環境で自由生活を営むもの、他の生物の体内に寄生するものなど、さまざまです。まずは、手分けをして土壌や河川などからメタモナス類を採集・分離しました。そのうえで、私たちの研究室が独自に確立していた培養法や解析技術を駆使してmRNAを抽出し、配列を解読した後、稲垣先生がいる計算科学研究センターに持ち込んで片っ端から解析していきました。
―― 3人の役割分担は?
稲垣氏: トランスクリプトーム解析後の進化学的解析は、私が一手に引き受けました。私も真核生物の進化に興味があり、ミトコンドリアや葉緑体がどのようにして細胞内に取り込まれてオルガネラになったのか、その経緯を分子系統解析により検討しています。計算科学研究センターに所属していますが、日頃からウェットな実験も行っており、今回、メタモナス類の採集も行いました。
神川氏: 私は、mRNAの抽出と配列解読、データの解釈や研究者のとりまとめを担当しました。今回は5カ国以上の研究者が互いの知見や技術を持ち寄って進めたので、とりまとめに苦労しました。スカイプを利用したり、別のプロジェクトでカナダに滞在した際に共同研究者と打ち合わせしたりもしました。共同研究者たちも一生懸命データを出してくれたのですが、実際にはデータが出てからが大変でした。私も稲垣先生同様、ウェットな実験もドライな計算科学もやりますが、今回は実験部分を担当したことになりますね。
橋本氏: すでにお話したように、私はメタモナス類の調整や解析技術部分を担当しました。私たちの研究室では、長年にわたって土壌や水中などの嫌気環境にいるメタモナス類の収集と培養を行い、真核生物の初期進化についての検討を進めています。培養技術の他、バクテリア混在下でメタモナス類だけを高純度に取り出す独自技術、MRO由来の核酸やタンパク質を抽出・解析するノウハウを蓄積してきています。今回の成果は、こうした知見をフルに活用した結果といえます。
―― どのような結果が得られたのでしょう?
神川氏: 真核生物94種、約4万のアミノ酸座位からなる大規模データマトリクスを得ることができ、まずは、これらの分子系統解析を行うことでフォルニカータ生物群、パラアバサリア生物群、プレアクソスティラ生物群が単系統群であること、フォルニカータ生物群とパラバサリア生物群が近縁でその外側にプレアクソスティラ生物群が位置付けられることが分かりました(図4)。
第2に、典型的なミトコンドリアを持つ生物に見られる「ミトコンドリアへの輸送に関与するタンパク質」について、代表的な18種のメタモナス類を対象にした相同性解析からは、ごく一部を除き相同性を持つ配列は見られませんでした。このことから、MROsへの輸送に関する機構は、ミトコンドリアへの輸送機構に比べて大きく縮退、あるいは変化している可能性が示唆されました。
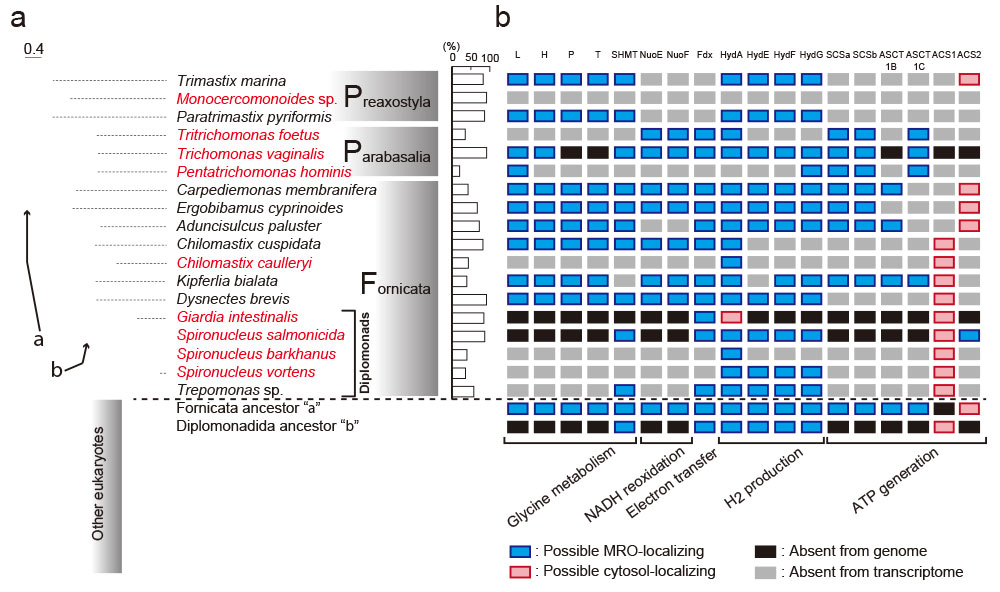
左側は、159タンパク質遺伝子、39,089アミノ酸座位、94生物種のデータに基づく最尤系統樹でメタモナス類を拡大表示している。右側の表で、■はMROへの局在が予測されること、■は細胞質への局在が予測されること、■はゲノムに存在しないこと、■は今回解析したトランスクリプトームには存在しないこと、をそれぞれ示している。 | 拡大する
第3に、同じ18種のメタモナス類を対象に、ミトコンドリアで見られるグリシン開裂系酵素4種類の相同性解析を行うと、自由生活性のものは4種類全てが保存されMROsで機能している可能性が高いことが分かりました。一方、寄生性のものは、その一部またはすべてを失っていました。寄生性のものは、宿主への依存により代謝系の一部をMROから捨て去る進化が起こったものと解釈できます(図4)。
第4に、嫌気的なATP合成能については、共通した仕組みがないと分かりました。細胞質内で合成するものもいれば、MROで合成するものもおり、合成に必要なタンパク質にも共通のルールはなく進化的に異なる酵素でした。つまり、嫌気環境が、ATP合成に関して幅広い多様性を許す環境であることが示唆されます(図4)。
最後に、MROの代謝系プロセスを検討した結果、メタモナス類進化のかなり早い段階でミトコンドリア縮退のイベントが起きたことがうかがえました。ジアルディアに至る系統ではATP合成能に加え、水素生成やセリン合成能も段階的に失われていき、その結果として、最も縮退したジアルディアのMRO代謝系では、鉄・硫黄クラスター合成系が残ったことが分かりました(図4、図5)。
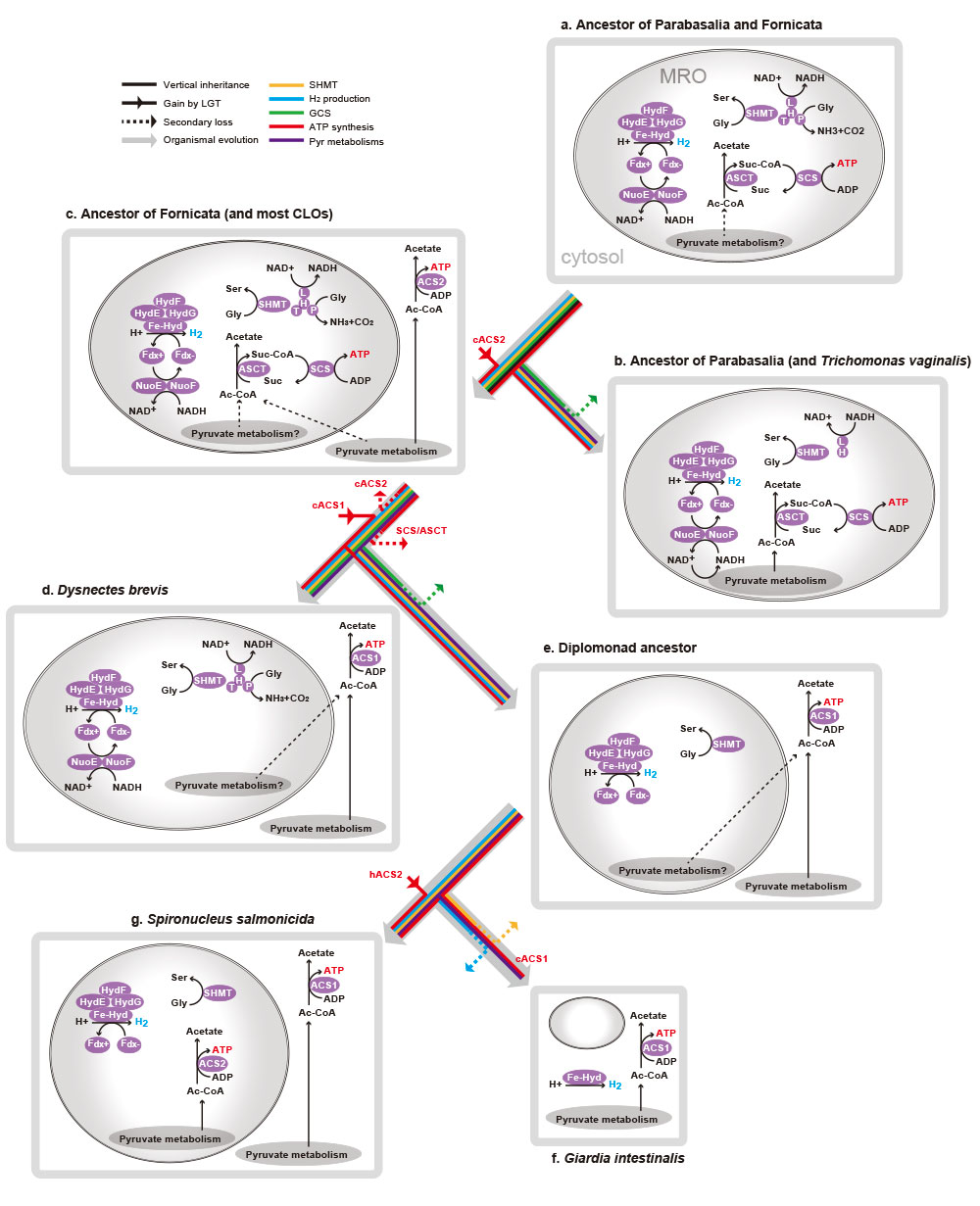
パラバサリアとフォルニカータの共通祖先におけるMROの代謝系が、系統樹のさまざまな進化段階を経てどのように縮退していったのかを模式的に示している。 | 拡大する
―― 多くの成果を挙げられましたが、残された課題はありますか?
稲垣氏: 今回の最大の成果は、トランスクリプトームの比較解析により、ミトコンドリアの機能が比較的残っているトリコモナスから、大半の機能を失ったジアルディアに至るまで、MROsの代謝機能の退縮経緯をきちんと推測できた点にあります。ただし、培養条件などにより検出できていないmRNAもあったはずで、今回で十分とはいえません。5カ国による共同研究はこれで終わりましたが、今後も協力関係を保ちつつ、別種でのトランスクリプトーム解析、既知データとの比較、MROsへの輸送に関与するタンパク質の検討などを進めていくことになると思います。
―― 今回の成果について、何らかの応用は考えられますか?
橋本氏: 近々の応用は難しいでしょう。可能性としては、病原性を持つメタモナス類について、今回見いだされたような「ヒトと異なる代謝システム」を薬剤標的にして創薬に生かせるかもしれません。ただ、すぐに応用できなくても、基礎研究の重要な知見を得たと自負しています。
神川氏: マングローブ林には自由生活性のメタモナス類が生息していますが、この先、さらに環境破壊が進んで具体的な保全策が必要になった際、メタモナス類を中心とする微生物環境や代謝についての知見が重要になってくるかもしれません。
稲垣氏: 環境中のDNA断片を集めて解読し、存在する微生物を特定するメタゲノム解析が注目されていますが、今回の成果は、メタゲノム解析では拾えない微生物がたくさん残されている点も示唆したと思います。シーケンス至上主義に陥りがちですが、今回、ラボで継代培養し、きちんと試料を用意して解析を進めることの価値も示せたのではないかと思います。
―― Nature Ecology & Evolution に投稿された経緯は?
神川氏: 初めはNature Microbiology に投稿したのですが、うまくいきませんでした。どうしようか考えた末に、私たちの成果には進化的な側面もあったので新創刊のNature Ecology & Evolution への投稿を思いつきました。Nature Ecology & Evolution がどのようなジャーナルなのか、興味もありました。投稿してみると、査読者がとてもポジティブで、編集者ともうまく進めることができました。いくつかの追加実験はしましたが、苦になるほどではありませんでした。研究者は進化的な議論と生態学的な議論を切り離してしまう傾向にありますが、今後はNature Ecology & Evolution が両者の融合に寄与するのではないかと期待しています。
橋本氏: Nature 関連誌に、こうした進化領域のジャーナルができたことは非常にうれしいですね。もちろん、Nature 本誌にも生態や進化領域が多く掲載されることを期待しています。
稲垣氏: 非常に魅力的なジャーナルが創刊されたので、今後もトライしていきたいですね。
―― ありがとうございました。
聞き手は、西村尚子(サイエンスライター)。
Nature Ecology & Evolution 掲載論文

Article: TrichomonasのヒドロゲノソームとGiardiaのマイトソームの起源の手がかりとなる細胞小器官
Organelles that illuminate the origins of Trichomonas hydrogenosomes and Giardia mitosomes
Nature Ecology & Evolution 1 : 0092 doi:10.1038/s41559-017-0092 | Published online 13 March 2017
Author Profile
神川 龍馬(かみかわ りょうま)
京都大学大学院地球環境学堂 助教
同大学院人間・環境学研究科 助教(併任)
| 2009年 | 京都大学大学院農学研究科応用生物科学専攻修了 博士(農学) |
| 2009年 | 日本学術振興会特別研究員(筑波大学計算科学研究センター) |
| 2011年 | 筑波大学生命環境系 特任助教 |
| 2013年 | 京都大学大学院地球環境学堂 助教(現職) 同大学大学院人間・環境学研究科 助教(併任・現職) |
| 2014年 | ダルハウジー大学(カナダ)客員研究員 |

稲垣 祐司(いながき ゆうじ)
筑波大学計算科学研究センター 教授
| 1995年 | 名古屋大学大学院理学研究科生物学専攻修了 博士(理学) |
| 1996年 | 日本学術振興会特別研究員(名古屋大学大学院理学研究科) |
| 1996年 | 株式会社JT生命誌研究館 奨励研究員 |
| 1998年 | 日本学術振興会海外特別研究員(カナダ・ダルハウジー大学) |
| 2000年 | ダルハウジー大学(カナダ)博士研究員 |
| 2004年 | 長浜バイオ大学 バイオサイエンス学部バイオサイエンス学科 講師 |
| 2005年 | 筑波大学大学院生命環境科学研究科 助教授 |
| 2007年 | 同 准教授 |
| 2011年 | 筑波大学生命環境系 准教授 |
| 2016年 | 同大学計算科学研究センター 教授(現職) |

橋本 哲男(はしもと てつお)
筑波大学生命環境系 教授
| 1988年 | 広島大学大学院生物圏科学研究科環境計画科学専攻修了 学術博士 |
| 1988年 | 広島大学原爆放射能医学研究所 助手 |
| 1991年 | 国立大学共同利用機関(現法人)統計数理研究所統計教育・情報センター 助手 |
| 1994年 | 同 助教授 |
| 1997年 | 同研究所調査実験解析研究系 助教授 |
| 2002年 | 同研究所予測制御研究系 助教授 |
| 2003年 | 筑波大学生物科学系 教授 |
| 2004年 | 同大学大学院生命環境科学研究科 教授 |
| 2011年 | 同大学生命環境系 教授(現職) |
