الوصفة التحريرية
لأبحاث أكثر استشهادًا
22 April 2025
نشرت بتاريخ 31 مايو 2023
عبر دراسة أُجريت على خلايا جذعية لذكور فئران، نجح فريق بحثي في تحويل خلايا تحتوي على مشيج متباين يشكله مزيج الكروموسومات الذكري X وY، إلى خلايا تحتوي على مشيج متجانس يشكله مزيج الكروموسومات الأنثوي الذي يتألف من اثنين من الكروموسوم X. وتستغل الدراسة ميل هذه الخلايا لاكتساب كروموسومات أو فقدها في أثناء الاستزراع الخلوي، بحيث يتولَد من التمايز المتتالي لهذه الخلايا بويضات قادرة على أداء وظائفها بصورة طبيعية، وينتج نسل حي جديد.
إن قدرة الخلايا الجذعية متعددة القدرات على تجديد نفسها وعلاج تلفها، والتمايز إلى أي من أنواع خلايا الجسم، يتيح فرصًا واعدة لدراسة الأمراض في بيئات الخلايا المستزرعة، وإنتاج خلايا علاجية متخصصة. غير أن إنبات الخلايا الجذعية مختبريًا له مخاطره أيضًا، لأن استزراعها لآماد زمنية طويلة قد يسفر عن عيوب بها، من أمثلتها اختلال الصيغة الصبغية، وهو خلل تتضاعف فيه الكروموسومات أو تُفقد كليًا1. وفي دراسة نُشرت مؤخرًا على صفحات دورية Nature، أفاد الباحث كِنتا موراكامي وفريقه البحثي2، بأن هذا النزوع إلى اختلال الصيغة الصبغية يمكن استغلاله لتحويل نوع من الخلايا متعددة القدرات، وهو الخلايا الجذعية الجنينية (mESCs)، من خلايا ذكرية إلى خلايا أنثوية بين فئران. ومن هنا، وجَّه الباحثون تلك الخلايا المتحولة جنسيًّا إلى تكوين بويضات يمكن تخصيبها لإنتاج نسل جديد، في إنجاز يُعدُّ لحظة فارقة في مجال البيولوجيا الإنجابية.
يبدأ إنتاج البويضات والحيوانات المنوية في أجنة الفئران، من طلائع أو خلايا سالفة تُعرف باسم «الخلايا الجنسية البدئية» (PGCs). والتمايز المبدئي لهذه الخلايا، الذي ينتج عنه بويضة أو حيوان منوي، لا يعتمد على كروموسومات جنسية طبيعية، وإنما على إشارات كيميائية محددة للنوع الجنسي، تصدر عن الغدة التناسلية المحيطة. ومع ذلك، فإن المراحل النهائية لتمايُز هذه الخلايا إلى بويضات مكتملة النمو، تتطلب بالفعل اثنان من الكروموسوم الجنسي X، في حين يتطلب تمايُزها إلى حيوانات منوية مكتملة النمو الكروموسومات الجنسية X وY3.
ويمكن أيضًا إنتاج البويضات والحيوانات المنوية للفئران مختبريًا، من خلاياها الجذعية الجنينية 5،4. ففي البداية ينشأ من هذه الخلايا خلايا تسمى «شبيهة الخلايا الجنسية البدئية» (PGC-like). أما الخطوة التالية، فهي استزراع هذه الخلية مع خلايا مأخوذة من مبيض أو خصية جنينيين (أو ما يكافئ ذلك من خلايا مولَّدة مختبريًّا)6، وذلك لبدء التمايز. ومن ثم، تظهر الحاجة إلى كروموسومات جنسية طبيعية، بُغية إتمام التمايز إلى بويضة أو حيوان منوي مكتملي النمو3.
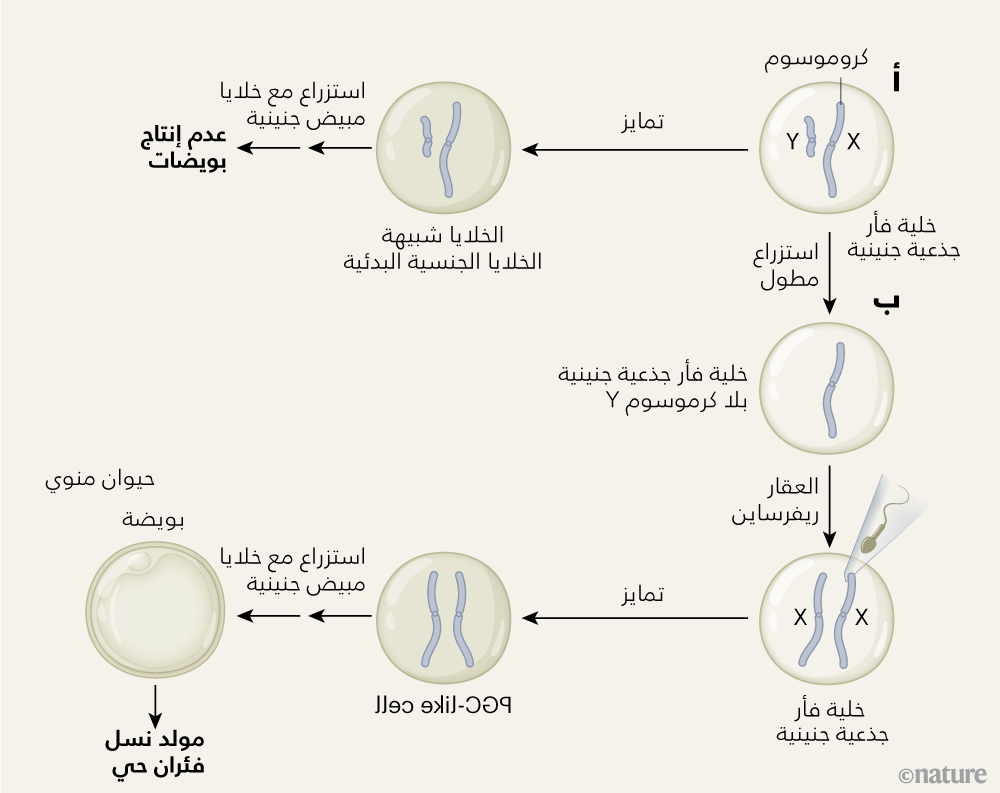
وفي هذه الدراسة التي بين أيدينا، سعى الباحث موراكامي وفريقه البحثي إلى توليد بويضات من خلايا ذكرية؛ وهو إنجاز من شأنه أن يتيح فرصًا هائلة لابتكار علاجات مستقبلية معززة للخصوبة. وبما أن إنتاج بويضات مكتملة النمو يتطلب وجود خلايا ذات مشيج متجانس، أي يحتوي على اثنان من الكروموسوم X، فقد ابتكر الفريق البحثي هنا طريقة للحصول على خلايا جذعية ذكرية نادرة، تلفظ الكروموسوم Y في المشيج المتباين بها، ثم تضاعف الكروموسوم X (الشكل 1).
----------------------------------------------
الشكل 1| بروتوكول لإنتاج بويضات من خلايا ذكرية. أ، يمكن للخلايا الجذعية الجنينية لفأر (mESCs) تحتوي على الكروموسومين X وY، أن تُستنبت مختبريًا، لتتحول إلى خلايا تُعرف باسم «شبيهة الخلايا الجنسية البدئية» (PGC-like)، وهي خلايا سالفة لخلايا البويضات والحيوانات المنوية. بيد أن هذا النوع الناشئ من الخلايا المحتوية على الكروموسومين X وY، لا يمكنه أن ينمو لينتج بويضة مكتملة النمو، حتى إذا استزرع معه خلايا مأخوذة من مبيض فأر جنيني. ب، أخضع موراكامي وفريقه البحثي2 الخلايا الجذعية الجنينية لفأر ذكر، للاستزراع لآماد زمنية طويلة، وهو ما أدى إلى فقد حوالي 6% منها للكروموسوم Y. ثم استزرع الفريق البحثي الخلايا التي فقدت الكروموسوم Y باستخدام عقار كيميائي يُسمى «ريفرساين» reversine، يعرقل آليات ضبط جودة انقسام الخلايا، بحيث يعزز احتمالية تضاعف الكروموسوم X. بعد ذلك، أخذ الفريق البحثي الخلايا الجذعية الجنينية الناتجة المحتوية على مشيج متجانس من اثنين من الكروموسوم X، وحثها على التمايز إلى الخلايا «شبيهة الخلايا الجنسية البدئية». وباستزراع تلك الخلايا مع خلايا مبيض جنينية، حفز ذلك نموها لتنتج بويضات قادرة على أداء وظائفها (لا تظهر في الشكل الكروموسومات)، ويمكن تخصيبها وزرعها في رحم إناث فئران، لإنتاج نسل فئران يافعة حية جديد.
----------------------------------------------
وقد صمم الفريق البحثي الخلايا الجذعية الجنينية للفئران الذكور التي استخدمها في تجربته، بحيث تحمل جينًا يشفِّر بروتينًا متألقًا باللون الأحمر، يوجد في الكروموسوم X لدى الفئران ويُشار له بالاختصار dsRED، وهو ما يسمح بمراقبة عدد هذا الكروموسوم في كل خلية. بعد ذلك، استزرع الفريق البحثي هذه الخلايا الجذعية الجنينية الذكرية، في ثماني جولات من إنباتها وإعادة واستنباتها في أطباق استزراع جديدة، وهي ظروف جعلت هذه الخلايا عرضة لفقد الكروموسوم Y على مرات متقطعة 1. ويمكن بسهولة رصد فقد أو حذف هذا الكروموسوم، عن طريق تقنية لتضخيم الحمض النووي، تُعرف باسم «تفاعل البلمرة المتسلسل» ويُشار لها بالرمز PCR. ووجد الفريق البحثي في الدراسة أن 6% من الخلايا فقدت كروموسومات Y التي تملكها، مكوِّنةً خلايا جذعية جنينية تحتوي على كروموسوم جنسي واحد هو الكروموسوم X (وقد أُطلق على هذه الخلايا الجذعية الجنينية، خلايا XO).
استمر موراكامي وفريقه البحثي في إنبات الخلايا XO، مع إجراء فحوص للكشف عن تضاعف الكروموسوم X الناتج عن استزراعها. ويحدث هذا التضاعف بسبب انقسام خلوي معيب؛ فبعد تنسُخ الحمض النووي، عادةً ما تنفصل أزواج الكروموسومX عن بعضها، مكوِّنة أزواجًا من الخلايا الابنة المتطابقة. غير أن حدوث خطأ في هذه العملية، قد يسفر عن انقسام خلوي، توجد فيه أزواج الكروموسوم X الناشئة عن الخلايا XO في خلية واحدة ابنة واحدة فقط، وبذلك تتكون خلية تحتوي على مشيج متجانس، أي على اثنين من الكروموسوم X. وقد استعان الفريق البحثي بتقنية لقياس شدة تألق البروتين الفلوري الأحمر، لتمييز الخلايا XO، التي وُجدت أول الاستزراع، عن هذه الخلايا الناتجة ذات المشيج المتجانس. ووجدوا أن معالجة خلايا XO بعقار يُسمى «ريفرساين» Reversine، من شأنه إلغاء نقطة تفتيش رئيسية في عملية الانقسام الخلوي، ليرتفع عدد مرات تضاعف الكروموسوم X.
بعد ذلك، عمد الفريق البحثي إلى جعل خلايا الفئران الجذعية الجنينية XX تتمايز إلى الخلايا «شبيهة الخلايا الجنسية البدئية»، وذلك باستخدام بروتوكول ابتُكر في وقت سابق، ثم أدخل الفريق البحثي إلى بيئة الاستزراع خلايا مأخوذة من مبيض فأر جنيني، وهو ما أصدر إشارات للخلايا «شبيهة الخلايا الجنسية البدئية»، لحثها على التمايُز إلى بويضات، حيث أتاح عندئذ المشيج المتجانس المؤلف من كروموسومي X اكتمال نشأة هذه البويضات. أما الخطوة الأخيرة، فهي تخصيب البويضات الناتجة مختبريًا، باستخدام حيوانات منوية برية النمط، وهو ما أنتج أجنة يمكن نقلها إلى رحم إناث الفئران، ومن ثم الحصول على نسل جديد قادر على البقاء. وأثبت الفريق البحثي أنه يمكن تحقيق نتائج مماثلة، عن طريق تحفيز حدوث تحول جنسي في «الخلايا الجذعية المستحثَّة متعددة القدرات» (iPSC)، وهي نوع من الخلايا متعددة القدرات المأخوذة من خلية أُخضعت للتمايز، وهي في هذه الدراسة خلية XY من جلد فأر، بحيث يحدث تحفيز التحول الجنسي عبر إعادة برمجة الهوية الخلوية لهذه الخلايا، لتصبح مشابهة للخلايا الجنينية، باستخدام عوامل نسخ.
جدير بالذكر أن حالات اختلال الصيغة الصبغية لا ترتبط كلها بالكروموسومات الجنسية. فعلى سبيل المثال، يؤدي ظهور نسخة إضافية من الكروموسوم الحادي والعشرين، في الحالة المعروفة باسم «تثلُّث الصِّبغي 21»، إلى ظهور متلازمة داون في البشر، وتتمثل بعض المظاهر المشابهة لهذه المتلازمة في الفئران، نتيجة لـ«تثلُّث الصِّبغي 16». وفي تجربة أخيرة أجراها موراكامي وفريقه البحثي، أوضحوا أن الخلايا الجذعية الجنينية للفئران، التي يشوبها «تثلُّث الصِّبغي 16»، لا يمكن أن تنمو منها بويضات مكتملة النمو. رغم ذلك، فمعالجة تلك الخلايا الجذعية الجنينية المحتوية على «التثلُّث الصِّبغي 16»، بعقار «ريفرساين»، من شأنها أن تحفز فقد الكروموسوم السادس عشر الإضافي، دون التأثير على باقي الكروموسومات. ونتيجة لذلك، استعادت الخلايا قدرتها على النمو لتشكِّل بويضات مكتملة النمو.
إن هذا البروتوكول الذي أعده موراكامي وفريقه البحثي، يفتح آفاقًا جديدة في مجال البيولوجيا الإنجابية وأبحاث الخصوبة. فالقدرة على توليد كل من البويضات والحيوانات المنوية، وبالتالي إنتاج نسل جديد من خلية جلدية ذات مشيج متباين تحتوي على الكروموسومين X وY، يمكن استخدامها في إنتاج سلالات من الفئران غير الهجينة، والتي من شأنها أن تتيح للتجارب المعملية بيئة جينية ثابتة، كما في حال التوائم المتطابقة. علاوة على ذلك، فاستراتيجية الباحثين في الكشف عن الخلايا التي فقدت كروموسومات، يمكن استخدامها لمعرفة السبب وراء ميل بعض الكروموسومات بدرجة أكبر من غيرها للإسهام في عمليات الانقسام الخلوي المعيب، سواء في الخلايا الجذعية الجنينية، أو في الخلايا السرطانية. على سبيل المثال، يمكن لأجزاء معينة من البِنى الكروموسومية (مثل موضع منها يُسمى القُسيم المركزي) أن تؤثر على احتمالية حدوث عمليات الانقسام الخلوي المعيب7، لكن يظل من غير المعروف، ما إذا كانت هذه الاحتمالية تعتمد على نوع الخلية، أم لا، وهو لغز يظل بحاجة إلى اكتشاف إجابته.
كذلك فإن هذه الدراسة قد توفر لنا آلية تسمح بتكاثر الثدييات المهددة بالانقراض، بالاعتماد على ذكر واحد منها، شريطة أن أمًّا بديلة من النوع نفسه أو من نوع ذي قرابة، يمكنها حمل الأجنة الناتجة حتى اكتمال نموها، بل إن الدراسة من شأنها أيضًا أن تتيح نموذجًا يسمح لمزيد من الفئات البشرية بإنجاب أطفال بيولوجيين، بما في ذلك الأزواج من الذكور مثليي الجنس، والمتحولين جنسيًّا، ومن يمتلكون كروموسومات XXY، مع تلافي الإشكاليات الأخلاقية والقانونية التي ينطوي عليها التبرع بالبويضات.
ورغم كل ما سبق، فإن البروتوكول الذي ابتكره الباحثون تشوبه أوجه قصور، أبرزها انخفاض كفاءته. فرغم أن حوالي 30% من خلايا الفئران الجذعية الجنينية المتحولة، نمت لتشكل بويضات مكتملة النمو، وأن 40% من تلك البويضات أمكن تخصيبها، فإن 1% فقط من الأجنة التي نُقلت إلى رحم أمٍّ بديلة، نمت ووُلدت حية. ويبقى أن نتيقن مما إذا كان القصور الذي أدى إلى انخفاض الكفاءة الإنتاجية لها البروتوكول، هو قصور تقني أم بيولوجي.
وفوق كل شيء، من غير المعروف حتى الآن، ما إذا كان البروتوكول سينجح في حال الخلايا الجذعية البشرية. فرغم نجاح تمايُز الخلايا الجذعية البشرية متعددة القدرات في هذا البروتوكول، إلى الخلايا «شبيهة الخلايا الجنسية البدئية»، يظل التمايز الجنسي المتتابع لإنتاج البويضات، يشوبه قصور، كما أنه يفتقر إلى الكفاءة الإنتاجية8، إذ إنه سيتطلب استخدام خلايا مبيض جنينية بشرية، وهي خلايا لم يثبت إلا مؤخرًا إمكان اشتقاقها مختبريًا من خلايا جذعية متعددة القدرات9.
بالإضافة إلى ذلك، رغم المساعي الجادة التي بذلها الفريق البحثي، لم يتمكن من إنتاج أجنة قابلة للحياة، من حيوانات منوية وبويضات مستنبتة مختبريًا من «خلايا جذعية مستحثة متعددة القدرات» مأخوذة من الذكر نفسه، وهو ما قد يتبين أنه يشكل عقبة كبيرة تعترض تحقيق الطموحات الموضحة في هذا المقال. ويكمن أحد التفسيرات المحتملة لذلك في البصمات الجينية؛ وهي إشارات كيميائية مُحددة للنوع الجنسي، تعدِّل الحمض النووي، ويرثها الجنين من أحد الأبوين. وتضمن البصمة الجينية التعبير في الجنين عن نسخ دون غيرها من جينات معينة، موروثة تحديدًا من الأم أو الأب، وهو ما يحول دون حدوث ظاهرة فرط التعبير الجيني. ومن شأن حدوث خطأ في نسب البصمات الجينية لكل من الأم والأب، أن يؤدي إلى حدوث فرط التعبير الجيني أو نقصه بشدة، وهو ما قد يتسبب في قصور في النمو10،11.
وبناء على ذلك، تُعد ظاهرة اختلال الصيغة الصبغية في الاستزراع الخلوي الروتيني، سلاحًا مزدوج المنفعة. فهي من ناحية تدعم خلق ظروف استزراع محسَّنة، لا سيما لإنتاج علاجات خلوية. ومن جانب آخر، تتيح فهمًا متعمقًا لعمليات خلوية أساسية. فما رصده موراكامي وفريقه البحثي من ندرة نسبية في حالات فقد الكروموسومات، يشير إلى أن الخلايا الجذعية متعددة القدرات، تمتلك آليات فعالة لإصلاح الحمض النووي وحراسة الجينوم، بغرض حماية الخلايا من الطفرات الضارة. ويكون سير هذه الآليات التي ما زالت مجهولة لنا، أكثر إحكامًا بدرجة كبيرة داخل الكائن الحي. ففي أجنة الفئران والبشر، تتحول الخلايا مختلة الصيغة الصبغية إلى سلالات خلوية في البِنى الداعمة المحيطة بالجنين، مثل المشيمة، لتوفر الحماية اللازمة له 12،1-14. وهذا التباين في السلوك، بين الخلايا في المختبر والخلايا داخل الكائن الحي، يشير إلى أنه ما زال أمامنا الكثير لنتعلمه، قبل أن نستطيع استخدام الخلايا الجذعية المستزرعة في إنتاج بويضات بشرية مختبريًا.
جوناثان باييرل وديانا جيه. ليرد، يعملان في قسم التوليد وأمراض النساء وعلوم الإنجاب، وفي مركز إيلي وإيديث برود للطب التجديدي وأبحاث الخلايا الجذعيّة، بجامعة كاليفورنيا، سان فرانسيسكو، سان فرانسيسكو، كاليفورنيا 94143، الولايات المتحدة الأمريكية.
البريد الإلكتروني: diana.laird@ucsf.edu
أقرَّ المؤلفان بعدم وجود تضارب في المصالح.
نُشِرت النسخة الإنجليزية من هذا المقال على الإنترنت في الخامس عشر من مارس عام 2023.
doi:10.1038/nmiddleeast.2023.170
1. Eggan, K. et al. Nature Biotechnol. 20, 455–459 (2002).
2. Murakami, K. et al. Nature 615, 900–906 (2023).
3. Taketo, T. Asian J. Androl. 17, 360–366 (2015).
4. Hayashi, K., Hikabe, O., Obata, Y. & Hirao, Y. Nature Protoc. 12, 1733–1744 (2017).
5. Hikabe, O. et al. Nature 539, 299–303 (2016).
6. Yoshino, T. et al. Science 373, eabe0237 (2021).
7. Worrall, J. T. et al. Cell Rep. 23, 3366–3380 (2018).
8. Yamashiro, C. et al. Science 362, 356–360 (2018).
9. Smela, M. D. P. et al. eLife 12, e83291 (2023).
10. Ferguson-Smith, A. C. & Bourc’his, D. eLife 7, e42368 (2018).
11. SanMiguel, J. M. & Bartolomei, M. S. Biol. Reprod. 99, 252–262 (2018).
12. Eakin, G. S., Hadjantonakis, A.-K., Papaioannou, V. E. & Behringer, R. R. Dev. Biol. 288, 150–159 (2005).
13. Frade, J. et al. Sci. Adv. 5, eaax4199 (2019).
14. Yang, M. et al. Nature Cell Biol. 23, 314–321 (2021).
الوصفة التحريرية لأبحاث أكثر استشهادًا
22 April 2025
20 April 2025
من عينة دم واحدة: الذكاء الاصطناعي يشخص السكري والإيدز وكوفيد
17 April 2025
اشترك للحصول على النشرة الإلكترونية المحدّثة كل أسبوعين لكي تبقى على اطلاع بكافة المستجدات على الموقع.
التسجيل في خدمة النشرات الإلكترونية (بالإنجليزية)
تواصل معنا: