حظر أستراليا غير المسبوق لوسائل التواصل الاجتماعي: تجربة حية للعلماء
11 January 2026
نشرت بتاريخ 18 أبريل 2022
تبين أن الناقل العصبي المعروف باسم الدوبامين، يعمل كمؤشر يطلق عملية التعلم في مركز الملاحة بدماغ ذبابة الفاكهة. ويتوقف معدل تعلم الذبابة للطرق على حركتها الدورانية، إذ تُسجَّل خلال تلك الحركة المعلومات البصرية المفيدة فقط، وتستخدمها لتحديث الخرائط الذهنية.
تواجه الأجهزة العصبية في الحيوانات، معضلة الموازنة بين تخزين معلومات جديدة عن محيطها، وتحديث ما جرى تخزينه سابقًا من بيانات مُرمِّزة للمدخلات الحسية عن هذا العالم المحيط. ومما قد يساعد على تحقيق هذا التوازن، مواءمة التحكم في المرونة التكيفية للشبكات العصبية، على نحو يسمح للحيوان باكتساب المعلومات الجديدة التي قد تفيده فقط. فما الذي يؤشر على هذا النوع من المعلومات، أو يخدم كمؤشر على "وقت التعلُم"؟ في بحث نشر مؤخرًا في دورية Nature، تطرح إيفيت فيشر وفريقها البحثي1، حلا لهذا السؤال، من خلال دراسة الشبكة العصبية المُرمِّزة لاتجاه دوران رأس الجسم في ذبابة الفاكهة. إذ توصل الفريق البحثي إلى وجود علاقة بين تغيُر معدل التعلم، وإطلاق الناقل العصبي الجزيئي المعروف باسم الدوبامين، والذي يستحثه انعطاف الحشرة في أثناء حركتها.
فلكي يصل المرء إلى وجهته، من المفيد أن يحفظ الطريق الذي يجب أن يسلكه. وعندما تمضي ذبابة الفاكهة في رحلة نحو ثمرة موز، أو نحو موقع تضع فيه بيضها، فإنها تتعلم خريطة لوجهتها، اعتمادًا على معالم بصرية ومؤشرات على اتجاهها2. بعدئذ، يوائم دماغ الذبابة بيانات هذه "الخريطة التقديرية للوجهة" مع الاتجاه الذي تتخذه في حركتها للوصول إلى مُحفِّز حسي مرغوب، أو يعتمد على تلك الخريطة في تقدير سرعة حركة الذبابة، وهو ما يسمح للحشرة بتذكر طريقها.
ويرتكز عمل هذا النظام الملاحي على خلايا عصبية معروفة باسم الخلايا البوصلة (EPG)، وهي تعمل على تلقي المعلومات عن المعالم في طريق الرحلة، من خلايا عصبية بصرية، لتختزن خريطة تقديرية لوجهة الذبابة. إذ تنتظم هذه الخلايا البوصلة مكونة تركيبًا أشبه بالحلقة، يسمى الجسم الإهليجي، يعمل موضع يرتفع مستوى نشاط الخلايا العصبية منه على ترميز زاوية اتجاه الذبابة. ومع كل انعطاف تسلكه الذبابة في أثناء حركتها، ينتقل نشاط هذه الخلايا إلى جزء آخر من الحلقة، لتحديث بيانات الزاوية. وهذه العلاقة بين عملية وضع خريطة دقيقة لمحيط الذبابة وعملية تحديد زاوية اتجاهها، على غرار تحديد الاتجاه في البوصلة، عشوائية في آلياتها؛ أي لا ينشط جزء بعينه من الحلقة، كلما اتجهت الذبابة نحو الشَّمال، على سبيل المثال، وإنما تُرمِّز الحلقة خريطة ثابتة تختلف باختلاف المشهد البصري، وتقترن بمشهد بصري معين. فينشط الموضع نفسه من الحلقة، عندما تتوجه الحشرة نحو نقطة معينة مصادفةً هذا المشهد البصري تحديدًا.
وما يسمح بهذه العملية التي تمزج بين آلية عشوائية وأخرى ثابتة، هو مرونة التشابكات العصبية التي تمتد من الخلايا العصبية البصرية إلى الخلايا البوصلة. إذ يمكن أن تتغير قوة هذه التشابكات سريعًا، لتعيد تشكيل خريطة الوجهة4،3. ومن ثم، قد يتصور المرء أن مثل هذا النظام، الذي يرمِّز باستمرار معلومات مكانية، يكتسب أيضًا باستمرار معلومات عن عناصر الوسط المحيط. والاحتمال البديل هو أن عملية التعلم تلك ينحصر حدوثها في اللحظات التي تكون بها الإشارات الحسية مُحملة بمعلومات أعلى قيمة حول المشهد البصري.
ومن هنا، تقصت فيشر وفريقها البحثي هذه الاحتمالات، من خلال التساؤل عما إذا كان بإمكان مجموعة من الخلايا العصبية المنتجة للدوبامين، والمعروفة باسم خلايا ExR2؛ تغيير معدل تحديث قوة الوصلات العصبية (درجة التشابك العصبي) بين الخلايا العصبية البصرية، والخلايا البوصلة في الجسم الإهليجي (انظر الشكل 1). فعندما تفرز آليات تعزيز التعلُم الدوبامين حال مخالفة نتائج الأفعال لتوقعات الدماغ، يجري تحجيم مقدار المعطيات أو المدخلات البيانية التي تعمل على تحديث نموذجنا عن العالم، بحيث يجري تحديثه عند كونه خاطئًا فقط. بيد أن بناء خريطة ذهنية للوجهة ينبني على نوع مختلف من التعلم؛ إذ يعمد الدماغ إلى تحديث نموذجه عن العالم المحيط، بغض النظر عن كون هذا النموذج خاطئا أم غير خاطئ. وعليه، إذا كان هناك من دور للدوبامين في هذه العملية، فماذا عساه أن يكون؟
للإجابة عن هذا السؤال، عمد الفريق البحثي إلى هندسة ذباب فاكهة وراثيًا، بحيث تُعبر خلايا ExR2 عن بروتين يسمى مؤشر الكالسيوم المُرمَّز جينيًا، وهو بروتين يدل إنتاجه على مستوى نشاط الخلايا العصبية. بعد ذلك، عمد الفريق البحثي إلى مراقبة نشاط خلايا ExR2 بعناية في أثناء مشي الذباب علي سطح كرة معلقة في الهواء، في الظلام (وهو ما يعني عدم ورود مدخلات بصرية بيانية إلى دماغ الذبابة). ووجد الفريق البحثي أن نشاط خلايا ExR2 ارتبط ارتباطًا وثيقا بالتغيرات التي تحدث لحظة بلحظة، في سرعة دوران الذبابة؛ إذ تبين أن نشاط هذه الخلايا العصبية، يرتفع كلما ازدادت سرعة دوران الذبابة حول الكرة. وبضم هذه الاكتشافات إلى نتائج توصل إليها الفريق البحثي سابقا4، وجد أن إطلاق الدوبامين، الذي تحفزه إشارات عصبية تصدر مع دوران الذبابة وتغييرها لاتجاهها يغير معدلات التعلم، لينتج خريطة دقيقة لوجهة الذبابة. ويشير ذلك إلى أن تعَلُّم الذبابة لطريقها ليس نتاج عملية متصلة، وأن انعطافاتها تبعث للدماغ برسالة مفادها: "حان وقت التعلم"، وذلك عندما تسنح فرصة اكتساب معلومات بصرية أعلى قيمةً.
----------------------------------------------------------------------------------------------------------------------------
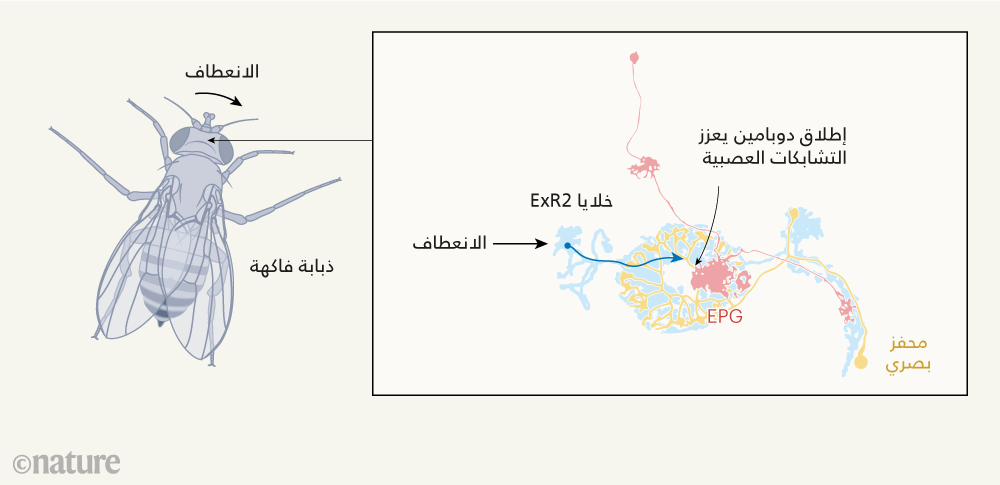
شكل 1. دور الناقل العصبي المعروف باسم الدوبامين في إرشاد ذباب الفاكهة إلى التوقيت الأمثل للتعلم. تعمل مجموعة معينة من الخلايا العصبية، تعرف باسم الخلايا البوصلة كجهاز بوصلة يُمكِّن ذباب الفاكهة من الوعي باتجاهه في مشهد ما. وقد وجدت فيشر وفريقها البحثي1، أن الإشارات العصبية التي تتولد لحظة انعطاف ذبابة الفاكهة، تُنَشِّط خلايا عصبية مُنتِجة للدوبامين، تعرف باسم ExR2. ويؤدي إطلاق الدوبامين الناجم عن هذا التنشيط، إلى تعزيز قوة الوصلات العصبية، أو ما يُعرف بالمشابك العصبية، بين الخلايا العصبية البصرية والخلايا البوصلة. وهو ما يؤدي بالتالي إلى ارتفاع معدل تعلُم الخلايا البوصلة. وبهذه الآلية، لا تستوعب هذه المنظومة الملاحية إلا البيانات البصرية عالية القيمة التي تساعد الذباب على تقدير وجهته. (تَظهَر الخلايا العصبية في الرسم على النحو الذي تنتظم به تشريحيًا مكونة خريطة دقيقة).
----------------------------------------------------------------------------------------------------------------------------
وللتحقق تجريبيًا مما إذا كان الدوبامين يتحكم في قوة التشابكات العصبية في هذه المنظومة، قاس الفريق البحثي أثر المحفزات البصرية على الخلايا البوصلة، مع التحكم في نشاط الخلايا العصبية ExR2. وسعيًا إلى القيام بذلك، لجأ هذه المرة إلى تقنية تسمى ملاقط فولطية الرقع الغشائية الراصدة للفسيولوجيا الكهربية، بهدف تسجيل النشاط الكهربي في خلايا بوصلة مُفردة، وحفَّز التعبير في الخلايا العصبية ExR2عن بروتين يوجد في قنوات الأيونات موجبة الشحنة التي يتحكم في منافذها ثلاثي أدينوسين الفوسفات، وهو ما أتاح للفريق البحثي بعدئذ التحكم في مستويات إطلاق الدوبامين من هذه الخلايا العصبية الأخيرة. وفي تجربة تستخدم منظومة افتراضية محاكية للواقع، دارت عارضة حول ذبابة مثبتة في مكانها. وتسبب هذا المُحفز البصري في إطلاق استجابات غير منتظمة من الخلايا العصبية البوصلة. غير أنه بعد تحفيز خلايا ExR2 لفترة قصيرة، أصبحت أنماط استجابة الخلايا البوصلة لهذا المحفز البصري أكثر ثباتًا. وهو ما يدل على أن خلايا ExR2 تُعزز تأثير المعطيات البصرية. وتأكد الفريق البحثي بعد ذلك، من خلال تصوير نشاط مجموعة الخلايا البوصلة، أن الخريطة الكاملة لمحيط الذبابة باختلاف اتجاه رأسها، أظهرت ارتباطًا واتساقًا أكبر مع موضع المحفز البصري بعد تنشيط خلايا ExR2.
وفي الخطوة الأخيرة من تجارب الفريق، للبرهنة على أهمية خلايا ExR2 العصبية في بناء خريطة دقيقة لوجهة الذبابة، ثَبَّطَ الفريق البحثي نشاط هذه الخلايا، عن طريق هندسة بعض الذباب وراثيًا، لينتج بروتينًا محددًا، من بروتينات قنوات البوتاسيوم. وفي هذه التجربة، اختبر الفريق البحثي سلوك الذباب مجددًا في منظومة افتراضية محاكية للواقع، لكنه هذه المرة أتاح للذباب إمكانية الحركة بحيث يتحرك المحفز البصري حوله مع حركته. وفي مجموعة المقارنة التي لم تُثبط فيها خلايا ExR2، أثبت هذا وجود ارتباط قوي بين اتجاه المحفز البصري وعملية تقدير اتجاه الذباب في الخلايا البوصلة. وظل هذا الارتباط قائمًا لعدة دقائق، حتى بعد أن جعل الفريق البحثي موضع المحفز البصري مغايرًا لاتجاه دوران الذبابة. أما في المجموعة التجريبية، التي جرى فيها تثبيط نشاط الخلايا العصبية ExR2، فقد صار هذا الارتباط أضعف؛ وهو ما دل على أن هذا النوع من الخلايا العصبية ضروري لاستحداث خريطة دقيقة لوجهة الذبابة، اعتمادًا على ما تقع عليه عينا الحشرة من معالم بصرية.
وقد ثبت منذ وقت طويل الدور الذي يضطلع به الدوبامين في تعزيز التعلم. بيد أن ما تلفت إليه فيشر وفريقها في هذا العمل البحثي هو أن هذا الناقل العصبي الجزيئي يتحكم أيضًا في معدل التعلم القائم على الاستكشاف؛ حيث ينتج المتغير السلوكي (معدل الانعطاف) في هذه الحالة إشارة تعمل من خلال إطلاقها للدوبامين على اختصار المعلومات الحسية الواردة، بحيث تُنتقى المعلومات الأعلى قيمة فقط. وهو ما يوضحه الفريق البحثي بالإشارة إلى أنه عند سير الذباب في خط مستقيم، يصبح حجم معلوماته البصرية محدودًا. ومن ثم، فإن الصورة الذهنية التي يكونها كنموذج عن العالم المحيط، بناءً على هذه المعلومات الجزئية، يرجح أن تكون غير دقيقة. ويعني هذا أن الجمع بين معدل تحديث بيانات الخلايا البوصلة ومعدل الانعطاف لا يحقق كفاءة في التعلم وحسب، على نحو يشبه ما يحدث عند عرض مقطع فيديو من تحديث بيانات البكسلات، بإضافة الجديد منها فقط الذي يختلف ما بين إطار عرض وآخر، لكنه أيضا إجراءٌ يضمن تجنب خطر المغالاة في تعويض نقص المعلومات.
بيد أن عدة أسئلة تبقى عالقة بلا إجابة، من أمثلتها ما الآلية التي يغير بها الدوبامين قوة الوصلات العصبية أو درجة التشابكات العصبية. على سبيل المثال، هل يحدث ذلك فقط من خلال تغيير عدد مستقبلات الناقل العصبي؟ أم أن هناك تغيرات بنيوية عصبية أخرى تسهم في عملية التعلم تلك؟ إضافة إلى ذلك، ما زال من غير الواضح، إلى أي مدى يمكن إطلاق الحكم بصحة هذا الاستنتاج القائم على نظرية إشارة "حان وقت التعلم" تلك. على سبيل المثال، إلى حد كبير، لم تتغير مراكز الملاحة في الدماغ بالحشرات مفصلية القدم على مدى تاريخها التطوري. غير أن استراتيجيات الحركة والتنقل تتعدد وتختلف بين أنواع هذه الحشرات، فكيف تتكيف الدوائر العصبية مع هذه الاختلافات؟ وفي الثدييات، تضطلع بالملاحة منظومة مشابهة وظيفيًا لكن مختلفة تركيبيًا عن تلك الموجودة في الحشرات، يُحتمل أنها تَستخدِم إشارات حركية مختلفة، ونواقل عصبية أخرى، لبناء الأسس المنطقية التي يقوم التعلم عليها لديها. فالمنظومة الملاحية لدى الثدييات، لا تختزن خرائط مكانية فحسب، بل تستطيع أيضًا بناء خرائط إدراكية تنبني بدرجة أكبر على مفاهيم مجردة6،5. فكيف يعثر الدماغ على الإشارة المناسبة التي تقوم بمقام إشارة «حان وقت التعلم»، عندما يريد تعلم هذه الخرائط؟ ومن شأن الإجابة عن هذه الأسئلة أن تلهم تصميم أنظمة ذكاء اصطناعي، قادرة على اختيار البيانات ذات الأهمية في ما تؤديه من مهام، لحظة بلحظة.
إستيفان تايز، وجريجوري إس. إكس. إي. جيفيريس، يعملان في قسم البيولوجيا العصبية، بمختبر البيولوجيا الجزيئية التابع لمجلس البحوث الطبية، بمدينة كيمبريدج CB2 0QH، بالمملكة المتحدة.
البريد الإلكتروني: itaisz@mrc-lmb.cam.ac.uk
jefferis@mrc-lmb.cam.ac.uk
doi:10.1038/nmiddleeast.2023.219
1. Fisher, Y. E., Marquis, M., D’Alessandro, I. & Wilson, R. I. Nature 612, 316–322 (2022).
2. Seelig, J. D. & Jayaraman, V. Nature 521, 186–191 (2015).
3. Fisher, Y. E., Lu, J., D’Alessandro, I. & Wilson, R. I. Nature 576, 121–125 (2019).
4. Kim, S. S., Hermundstad, A. M., Romani, S., Abbott, L. F. & Jayaraman, V. Nature 576, 126–131 (2019).
5. Constantinescu, A. O., O’Reilly, J. X. & Behrens, T. E. J. Science 352, 1464–1468 (2016).
6. Nieh, E. H. et al. Nature 595, 80–84 (2021).
11 January 2026
31 December 2025
30 December 2025
اشترك للحصول على النشرة الإلكترونية المحدّثة كل أسبوعين لكي تبقى على اطلاع بكافة المستجدات على الموقع.
التسجيل في خدمة النشرات الإلكترونية (بالإنجليزية)
تواصل معنا: